1. Introduction
Since the first radical mastectomy conducted by William Halsted in 1882 to treat breast cancer [1], cancer treatment and patient survival have been substantially improved [2]. However, the battle against cancer is by far not yet won. One of the central challenges for cancer treatment is identifying patients who are more likely to respond favorably to a given anticancer therapy. Clinical and pathological parameters routinely used for diagnosing, predicting outcomes, and treatment selection, often fail to describe tumor heterogeneity comprehensively. A consensus is that the combination of pathological and clinical parameters with biological prognosticators better explains the heterogeneity in the treatment outcomes. Identification of these biological stratifiers, as well as delivering more tailored treatment, is the ultimate goal of personalized and precise medicine. As the indicators of tumor presence and progression, cancer biomarkers are tightly connected to the functional capabilities of tumor cells, which Hanahan and Weinberg describe as cancer hallmarks [3,4]. These hallmarks of tumor functions include chronic proliferation and immortality, circumventing growth suppression and cell death, activating angiogenesis and metastasis, genome instability and epigenetic reprogramming, transitory senescence and immune evasion, tumor-inducing inflammation and tumor-promoting microbiota, cellular phenotypic plasticity and reprogrammed cellular metabolism [3,4]. As our knowledge of tumor mechanisms and complexity progressed, the physical parameters of cancer cells and their microenvironment gained appreciation as critical regulators of tumor progression and therapy resistance and potential prognostic traits associated with clinical outcomes in patients with different types of malignancies.
The physical traits of cancer include the characteristics of tumor cells, such as mechanical, thermal, electrical properties and the physical microenvironment, including solid and liquid pressure, tissue stiffness and architecture [5,6]. During tumor development, the physical abnormalities of tumors emerge as a consequence of biological hallmarks of cancer but also induce more aggressive functional tumor capabilities and result in a vicious cycle driving tumor progression. Thus, a comprehensive analysis of both biological and physical cancer parameters is critical for developing more robust prognostic stratifiers, discovering yet unexplored therapeutic targets and improving treatment efficiency. Similar to the biological hallmarks, physical abnormalities of tumors have a high inter- and intratumoral heterogeneity and can be dynamically changed during the natural tumor history and as a result of treatment. The heterogeneity of biophysical tumor characteristics and their clinical relevance highlight the necessity of their spatial and temporal analysis in cancer patients. This review focuses on the role of physical parameters of tumor cells and microenvironment for tumor development, progression and therapy resistance, and potential treatment strategies to target tumor physical traits. The analytic approaches for the assessment of these parameters and their potential implementation in clinical practice will also be discussed.
2. Mechanical Properties of Cancer Cells
2.1 Introduction to the topic
The idea that tissue mechanical properties can inform on certain tissue abnormalities, including neoplastic lesions, is as old as the practice of tissue palpation, where the differential mechanical properties of the probed tissues are revealed qualitatively [7]. Quantitative data can be obtained with higher sensitivity and reproducibility using more sophisticated physical tools that allow for in situ mapping of the tissue of interest, e.g., elastography techniques [8,9], or ex situ, e.g., by shear rheometry [10] and atomic force microscopy (AFM) [11]. Of relevance, these changes in mechanical properties are characteristic of the development and progression of tumors and can thereby reflect the tumor progression state. This is supported by previous reports showing correlations between tissue mechanical properties and breast tumor subtype and stage [12] or studies suggesting the value of tissue stiffness as diagnostic biomarkers e.g., for prostate cancer [13,14] and liver cancer [15]. In many cases, solid tumors are found to be stiffer than normal tissues. However, there are also studies where no significant differences between normal and tumor tissues can be seen, as in the case of cervical cancer [16].
Biological tissues are soft, complex, and heterogeneous materials composed of various cell types and extracellular matrix (ECM) that provide a scaffold to embedded cells. Mechanical changes in tumors span from subcellular to tissue scales and can be heterogeneous at all levels. At the tissue scale, they often arise from alterations in ECM architecture and mechanical characteristics. In some solid tumors, including breast, prostate and pancreatic cancer, a denser and more rigid collagen network is commonly seen, and this desmoplastic response can be associated with poorer patient outcomes [17–20]. Increased collagen deposition and cross-linking are driven by cancer-associated fibroblasts that are characterized by a more contractile phenotype [21,22]. Besides increased matrix stiffness, interstitial fluid pressure, solid stress, and cellular interactions can contribute to overall altered tissue mechanical properties. Interstitial fluid pressure, driven by blood and lymphatic fluids, differs between normal tissues and tumors [5,23]. Interstitial fluids can also flow due to pressure differences between blood and lymphatic systems, imposing wall shear stresses on surrounding cells and tissues [24,25]. A tissue mechanical parameter that appears relevant with regard to diagnostics is tissue fluidity, which is also accessible through elastography measurements. Tumor tissues can be more fluid-like or more solid-like, which is not simply related to the tissue’s water content but also to collective cell behaviors, e.g., cell-cell and cell-ECM interactions [26]. These collective cell behaviors can be physically described as jamming and unjamming transitions that arise also in non-biological systems [27,28]. Last but not least, the tumor constituent cells themselves can exhibit cell-to-cell heterogeneities but also internal heterogeneity in their biochemical composition, consisting of a liquid or gel phase interlaced with fibrillar networks building the cytoskeleton. Depending on the tissue, these properties can largely affect the tissue mechanical response as, for instance, recently shown for breast and cervix [16] and brain tumors [29]. Due to their structural complexity, it is expected that cells, both healthy and cancerous, display considerable variability in their viscoelastic properties - a phenomenon amply confirmed by experimental studies [6,30–32]. This variability depends in particular on the cell types, cell state (cell cycle, shape) and the spatial and time scale at which these properties are measured. A consensus has emerged defining three characteristic length scales in cell mechanics [32]: the intracellular scale (<10 μm), the whole-cell scale (~10–30 μm), and the tissue scale (>30 μm) [33].
Here, the focus will be on cell-scale mechanical changes and measurement techniques assessing apparent elastic and viscoelastic changes in cells at the intracellular and whole-cell scales. Specifically, a meta-analysis of previous studies that have embarked on comparative analyses of healthy and cancerous tissue and cells will be provided. Furthermore, the biological bases of viscoelastic alterations, commonly used tools to assess cell mechanical properties at different scales, will be described.
2.2 Essential terminology and definitions
In the field of cell biomechanics, physical quantities like stiffness, elastic modulus, and viscosity are widely used to describe the mechanical response of cells and tissues [34,35]. The stiffness coefficient (K) is defined as the ratio of the applied force to the resulting displacement at the point of application. K depends not only on the material elastic properties but also on the sample shape and dimensions. The elastic modulus, in contrast, is an intrinsic bulk property of the biological material, denoted G for shear and E for extension/compression and does not depend on geometry. In many biomechanical experiments, including AFM, it is often challenging to determine whether the measured value corresponds to the stiffness coefficient or the elastic modulus. The convention adopted by many research groups is to refer to these measurements as the apparent Young's modulus, EApp[30,33–35]. Additionally, certain techniques generate quantitative parameters, such as shear wave speed or Brillouin frequency shifts [36,37], that are indirect stiffness indicators, rather than physical quantities known from continuum mechanics or rheology. The above quantities, K, G, and E can also be measured as a function of frequency, leading to complex variables K*(ω), G*(ω), and E*(ω). The frequency dependence also allows the data to be compared with predictions from the constitutive equations of known viscoelastic models [38]. Active or passive microrheology using internalized micron-sized probes, in turn, measures the shear viscosity (η0) and elastic modulus (G) of the cytoplasm [39]. When experimental conditions are not at low-frequency and in shear mode, the terms apparent viscosity (ηApp) or modulus (GApp) are used instead.
2.3 A meta-analysis of comparative studies assessing cell mechanical properties
2.3.1 Cell cortical stiffness measurements
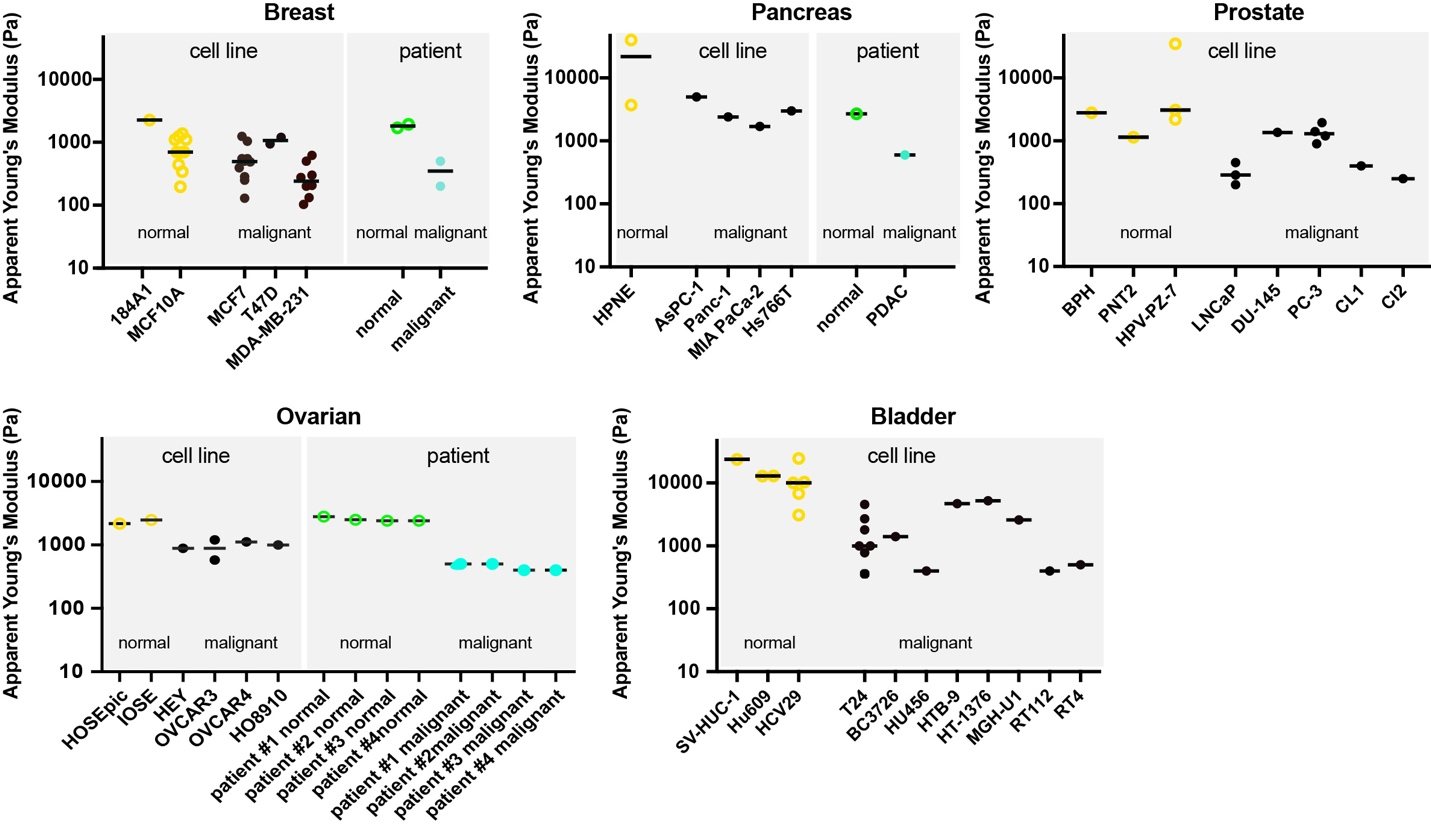
The first reports that cancer cells are softer than normal cells were published almost two decades ago [30,33,34,40,41]. Since then, multiple studies have been conducted on the mechanical analysis of established cancer cell lines and patient-derived cells. A meta-analysis on a broader collection of studies contrasting cancer and normal cell stiffness is presented in Figure 1. It comprises a total of 30 studies across six different cancer types: breast (n = 14), pancreas (n = 3), bladder (n = 8), prostate (n = 3), and ovarian (n = 2) cancer. Although there are many more published studies that report on differences depending on disease state, we focused here on studies reporting comparable parameters such as the apparent Young’s modulus EApp. Among the selected studies, AFM-based methods dominate, since AFM is by far the most commonly employed technique to assess cell mechanical properties. In Figure 1, apparent Young’s moduli are plotted for the abovementioned tumor types. Most studies focused on the assessment of established cancer cell lines, mostly breast cancer cells, and a high degree of redundancy becomes apparent. In many cases, even the same cell lines were measured by different research groups, suggesting a relatively high reproducibility of results. However, the limited number of studied models also limits the possibility of drawing more general conclusions. Of note, the prevalence of cancer cell lines compared to patient-derived material appears critical when cell mechanical parameters are to be evaluated with regard to their clinical relevance. The underlying reasons for using cell lines are obvious, since the use of clinically relevant samples is comparatively more challenging since it involves ethical approvals, collaborations with clinicians providing samples, and patient-to-patient heterogeneities, among other challenges. Nevertheless, it will be important to expand studies on patient-derived cells in the future.
In most cases, there is accordance that normal cells have higher apparent Young’s moduli compared to cancer cells. In some cases, the more invasive cell lines or metastatic primary cells had even lower stiffness values compared to less invasive cells. There are some exceptions, however, e.g., where increased cellular stiffness values are shown for invasive compared to low invasive prostate cancer cells [42]. Of note, EApp across studies widely ranges from approximately 100 to up to 30.000 Pa. Studies reporting higher values up to 100.000 Pa were excluded here, as they appear unrealistic for soft biological tissues. Generally, AFM indentation tests with sharp indenters on spread cells can result in higher values than measurements on rounded cells. In some cases, especially when spread cells with flat cellular extensions are tested, the underlying stiff substrate also might have affected the obtained values.
Although the viscoelastic nature of cells is well known [67,68], most cell mechanics surveys to date have been limited to an apparent elastic response (>80% of all studies), with only a small fraction addressing cytoplasmic viscosity properties. Here, we extended our literature search to studies that assess cell viscosity, aiming to explore its potential as a biomarker for cancer diagnostics.
2.3.2 Whole-cell viscosity
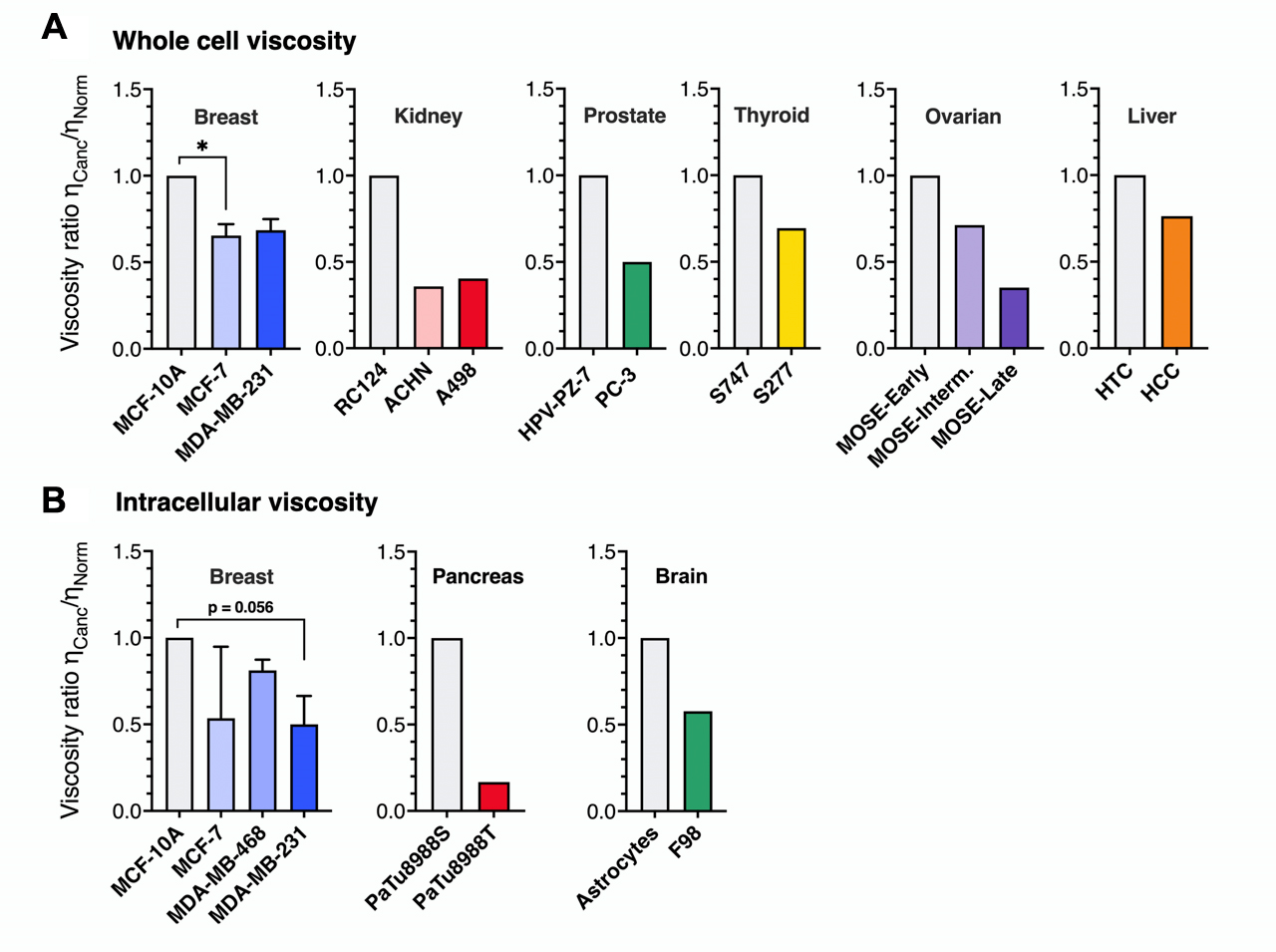
To measure the whole-cell apparent viscosity ηApp, researchers employ microrheology techniques inspired by stress relaxation or creep tests, commonly used in conventional rheology [38]. Our meta-analysis includes nine studies across six different cancer types: breast, kidney, prostate, thyroid, ovarian, and liver. Overall, the data reveal a strong preference for AFM-based methods [43–45,48,69–72]. Other techniques are micropipette aspiration [72] and micro-electromechanical systems (MEMS) resonant sensor [45]. Whole-cell viscosity measurements exhibited significant variability, with ηApp ranging from 3 to 500 Pa · s (3,000 to 500,000 times the viscosity of cell culture medium [73]. This variation is likely due to differences in the cell and cancer types studied, and in the case of AFM, also influenced by the specific protocols and tips employed [30,74]. The viscosity ratio ηCanc/ηNorm between cancerous and healthy cells was calculated for six cancer types to allow for comparison (Figure 2A). This figure consistently reveals a clear pattern: cancer cells exhibit lower viscosities than healthy cells. Overall, this reduction in viscosity averaged 44%, comparable to the decrease observed at the intracellular level (Figure 2B). For cancers with data on cells of increasing metastatic potential, only ovarian cancer cells (early, intermediate, and late MOSE cells) show a distinction between low and high invasiveness. This finding indicates that cancer cells undergo a fluidization of their mechanical properties compared to normal cells.
2.3.3 Cytoplasmic viscosity
To measure the cytoplasmic viscosity of healthy and cancerous cells, particle-tracking microrheology, optical tweezers, and magnetic rotational spectroscopy were applied across seven studies covering three cancer types: breast, brain, and pancreas [75–81]. In these techniques, micron or submicron-sized particles are dispersed in the cytosol, and their passive or active movements are tracked, allowing local viscosity measurement. Here again, breast cancer cells dominate the data, specifically four cell lines with increasing metastatic potential, MCF-10A, MCF-7, MDA-MB-468, and MDA-MB-231, respectively, accounting for 75% of all intracellular assays. For conventions, the static shear viscosity, such as that obtained in particle-tracking microrheology and magnetic rotational spectroscopy, will be noted η0 [38], and ηApp, will be used when the experimental conditions are not low-frequency shear.
According to literature data, particle-tracking data [78–81] show viscosity values ranging from 0.3 to 6 Pa · s – 300 to 6,000 times that of cell culture medium at physiological temperature [73], while active methods report significantly higher viscosities, between 10 and 50 Pa · s [75–77]. These data heterogeneities could be attributed to the size of the probes used, from the order of a hundred nanometers for the former and a micron for the latter [77]. To account for the differences in absolute values across techniques and to allow for comparison, we expressed existing intracellular results as the ratio: ηCanc/ηNorm, where ηCanc and ηNorm represent the viscosities of cancerous and normal cells, respectively. Figure 2A displays the ηCanc/ηNorm for breast, pancreas, and brain cancer types, with the first bar in each histogram set to 1 by definition. The figure highlights a consistent decrease in ηCanc/ηNorm across all cancer types, corresponding to an average 51% reduction in viscosity. This suggests a general trend toward increased cytoplasmic fluidization in cancerous cells.
For breast cells, Young’s moduli of MCF-7 and MDA-MB-231 tumorigenic lines were lower than those of healthy cells, but the difference was insufficient to differentiate between malignant and non-malignant cells clearly. Magnetic rotational spectroscopy did, however, reveal a significant difference, with MCF-7 cells showing five times higher intracellular viscosity than MDA-MB-231 [77]. This underscores cytoplasmic viscosity sensitivity to metastatic potential, suggesting that it could serve as a mechanical biomarker for cancer cells with high metastatic potential. The preceding analysis shows that findings on cancer cell viscosity have been insufficiently documented and that further research is required, especially by expanding the scope to include more diverse cell lines and cancer types. In the following sections, we will discuss the techniques utilized to measure the viscoelastic cell properties, which are summarized in Table 1.
2.4 Methodology
2.4.1 Atomic force microscopy (AFM)
Of all the mechanobiology techniques available today, AFM is by far the most widely used [30,34,41]. AFM is an ultrasensitive technique to measure forces down to the piconewton range [83,84]. It relies on a force sensor, the AFM cantilever that can be equipped with a nano- or macro-sized indenter (e.g., a pyramidal tip or also larger e.g., a colloidal probe in the range of 5–10 μm diameter). To probe cell mechanical properties, the cantilever is brought into contact with a cell (either round or in adherent state) with a pre-set force or indentation depth, and from the resultant force-distance curve the force-indentation relation can be derived. By fitting the force indentation curve to a model of choice (most commonly a Hertz model for simple indentation tests [85,86], an apparent elastic modulus EApp can be quantified, which represents the most commonly reported parameter for cell and tissue mechanical measurements. Alternative models include, for instance, the Oliver Pharr model [11], and the liquid droplet model [87]. In a configuration where the stress/strain response is measured over time, AFM can also be employed to determine the apparent viscosity of single cells [43,44,48,70,82]. Such measurements include stress relaxation [88], creep compliance, or oscillatory probing protocols [44,89,90]. In such cases, viscoelastic mechanical response models, such as the Kelvin-Voigt, Standard Linear Liquid or Standard Linear Solid models, or more complex rheological models can be employed to derive viscoelastic cell parameters from force versus time traces [33,89]. Alternatively, information on the viscoelastic properties can be obtained from the indentation part of the curve using viscoelastic relaxation models [91,92]. The strength of AFM lies in its ability to integrate high-resolution topographical imaging with force mapping, an essential configuration for biomechanical studies. AFM can also be combined with optical imaging or spectrometric techniques, e.g., advanced optical techniques such as confocal microscopy [93], total internal reflection fluorescence (TIRF) microscopy [93], calcium imaging [42], or Raman microscopy [94] to obtain additional information on cell state. While AFM is versatile with regards to cell shapes (both rounded and adherent), it is typically restricted to probing the cell surface, and furthermore relatively low in throughput (typically 100 cells/hour).
2.4.2 Micropipette aspiration
This technique applies negative pressure to draw a part of a single cell into a narrow glass pipet [95]. The living cell, suspended in solution, is first immobilized at the pipet tip before suction is applied. Using optical microscopy, the portion of the cell entering the pipet for a given pressure is monitored over time. The kinetic time course of cell deformation resembles creep experiments in rheology, where stress is applied and deformation is recorded over time [38]. Using viscoelastic models to fit the length time series, the cell viscoelastic properties, including its apparent viscosity ηApp, can be calculated [72].
2.4.3 Microfluidic techniques
To overcome the limitation of low throughput inherent to some aforementioned assays, microfluidic techniques can be attractive, e.g., for cell deformability cytometry [96–99], microconstriction arrays [100], or shear flow deformation cytometry [101] studies. These techniques have in common that suspended cells are passing through a microfluidic channel where they deform under fluid shear stress [99,101] or have to squeeze under pressure through narrow channel constrictions [100]. Resultant cell deformations or passage time are quantifiable parameters that can be assessed in timelapse videomicroscopy recordings. When the applied shear stresses are known, the viscoelastic properties of the cells can be inferred. High rates of deformation in the millisecond range (as in deformability cytometry) typically result in increased elastic moduli values compared to deformations in the second range [102]. Advantageously, these techniques can also be combined with fluorescent detection, which allows the mapping of mechanical parameters to specific markers [98]. Recent real-time deformation cytometry (RT-DC) based studies have revealed mechanical changes of transformed mammary cells [103,104], mechanical effects of different drugs on leukemic cells [100], and native and malignant cells in pleural fluids [99]. In addition, differences between normal and cancer cells were seen in cells derived from solid tumor tissues through digestion or mechanical dispersion [105].
2.4.4 Optical stretcher
Using the optical stretcher, cells are mechanically deformed in a contact-less manner using a dual-beam laser trap [106]. In a configuration combined with a microfluidic pump system, cells enter the optical flow chamber one by one, where they are trapped and stretched along the axis of the opposing laser beams [107]. The resultant cell elongation under stress is monitored over time and information about the cells’ viscoelastic properties of the cell can be quantitatively derived [108]. Using the optical stretcher, the mechanical properties of the breast [16,106], cervix [16], and brain cancer cells [26] were measured and compared to normal cells. Automated setups also provide the opportunity to assess the mechanical properties of hundreds or thousands of cells and to distinguish subpopulations to reveal cell mechanical heterogeneity [26,29].
2.4.5 Brillouin microscopy
While most of the above-mentioned techniques require physical contact between the probe and the material to be probed, optical elastography probes have the advantage that they can interrogate cells in a rather non-invasive manner. Brillouin scattering, discovered nearly a century ago, has recently been exploited for biomechanical measurements of biological specimens by Brillouin microscopy [36,109–112]. Brillouin microscopy measures the inelastic Brillouin scattering that arises from the interaction of photons with acoustic vibrations in a material. In most biomechanical studies, the Brillouin frequency shift, which is related to the longitudinal modulus, is used as a quantitative parameter to describe the elastic properties of biological specimens. Despite its non-invasiveness, an advantage of Brillouin microscopy is its high spatial resolution (in the subcellular range, depending on the optical setup), allowing for in situ 3D mapping of the mechanical properties of biological specimens. This technique can be employed on single cells but advantageously also on transparent tissues or multicellular structures such as organoids [36,111,113,114]. Calculation of a longitudinal modulus from the Brillouin frequency shift requires measurements of the refractive index, e.g., measured by optical diffraction tomography [115]. A limitation of the technique is that it is not possible to directly convert the longitudinal modulus at high frequency (GHz range) to shear moduli obtained by common probing techniques, e.g., AFM [110]. Nevertheless, previous studies have revealed correlations between results obtained by AFM and Brillouin, e.g., when osmotically perturbing cells and cell aggregates, and when applying drugs that interfere with major cellular structural elements, such as F-actin [113,116]. Differences between normal and cancer cells were also reported [36].
2.4.6 Particle-tracking microrheology
Particle-tracking microrheology has been a well-established method for several decades [39,95] to measure the static viscosity η0 in confined environments, particularly in living cells [117]. Particle-tracking microrheology employs high-speed optical microscopy to track the movement of particles in the 100 nm range embedded in the cytoplasm. To enhance particle detection and prevent their internalization into endosomes, which can lead to directed movement of the probes, fluorescent particles are used and introduced via ballistic injection [80,118]. The particle trajectories are analyzed in terms of the mean-squared displacement, and interpreted thanks to the Stokes-Einstein equation that relate the diffusion coefficient to η0 [119]. The technique is non-invasive, requires minimal material (<1 pL), and can be applied in both passive and active matter.
2.4.7 Optical tweezers
More recently, advanced methods such as optical tweezers [75,120] have expanded the range of intracellular rheological techniques for actively probing cytoplasmic viscous properties. This technique uses a fixed optical trap and microscope to study intracellular mechanics [120]. An infrared laser traps a micron-sized bead within the cytoplasm, allowing steady or oscillating manipulation. For viscosity measurement, the bead is displaced by moving the microscope stage, and the optical tweezers apply a spring-like force, causing the bead to relax back toward the trap center. This time-dependent relaxation is analyzed using the Standard Linear Liquid model [38] and allows simultaneous determination of the elastic modulus G0 and apparent viscosity ηApp of the intracellular medium. Experiments on breast [75] and brain [76] cancer cells reveal mitigated viscosity and elasticity properties compared to healthy ones.
2.4.8 Magnetic rotational spectroscopy (MRS)
This technique leverages the hydrodynamic instability found for anisotropic magnetic objects in a rotating magnetic field [121]. Berret et al. adapted MRS to intracellular environments using micron-sized magnetic wires that are spontaneously internalized in the cytosol and non-toxic to cells [122,123]. As the angular frequency ω increases, a transition from synchronous to asynchronous regime occurs at the critical frequency ωC, which varies inversely with the static viscosity η0. In viscoelastic media, the high-frequency wire oscillation amplitude varies inversely with the elastic modulus (G0). MRS hence enables the simultaneous measurement of local viscosity and elasticity, making it a powerful tool for probing local rheological properties of fluids or solids [124,125], including those of living cells.
2.5 Biological basis of cell mechanical alterations in cancer cells
The measured cellular response to external forces is dominated by the cytoskeleton, which is made up of dynamic and entangled polymer networks of F-actin, intermediate filaments and microtubules [126,127], and internal structures such as the nucleus [128], and other organelles [129]. Cancer progression is associated with remodeling of the actin cytoskeleton [89,104,130] and the microtubule network [49]. Also, changes in vimentin levels have been linked to cancer progression and even proposed as predictors of patient outcomes in lung carcinomas [131]. Recent studies have reported qualitative changes in cytoskeletal structures in normal and cancer cells that are associated with the measured changes in cell viscoelasticity [69,132]. Recently, induction of oncogene expression and epithelial-to-mesenchymal transition (EMT) has been demonstrated to directly affect cancer cell cytoskeleton and the mechanical properties of cancer cells [89,103,104].
To which extent the different cytoskeletal structural elements contribute to the cellular response to force during the measurement is also dependent on geometric features of the measurement technique (e.g., indenter size and shape), the probed cell’s shape, the time scale of the measurement technique (milliseconds to seconds), the cellular region being probed (nucleus versus lamellipodium, cytoplasmic or nuclear internal structures [50] and cell cycle stage [89,103,133]. For instance, AFM experiments are typically dominated by the actin cortex, which is located beneath the cell membrane [134]. Experiments perturbing the actin cytoskeleton (e.g., by cytochalasin D, latrunculin B) can have drastic effects when indenting the cell surface using an AFM cantilever [50] or when deforming suspended cells [97], albeit latter depends on the timescale: while cells deforming under moderate flow rates [97] are sensitive to actin-depolymerizing drugs, assays employing high rates were shown to be insensitive to these changes [99,102]. Similarly, interfering with actomyosin contractility (e.g., by blebbistatin) typically softens cells, at least when they are adherent to a surface where they form stress fibrils [89,135]. However, targeting myosin activity in suspended cells can even have the opposite, stiffening effect [136].
Also, the location where the cell is probed can matter, particularly in adherent cells. Previous studies have, for instance, revealed different mechanical properties when probing nuclear and perinuclear regions [50]. For thin regions probed by AFM, thin layer corrections should be taken into account [88,137]. Deeper indentations by an AFM tip can also reveal contributions of the intermediate filaments [138] or the nuclear lamina (along with F-actin depolymerization) [139]. Manipulations of microtubules can have opposing effects, where nocodazole can soften cells [75], have no effect, or even stiffen cells [140]. The response could be cell type dependent but also be related to crosstalk between microtubules and F-actin [126]. In some cases, e.g., for intermediate filaments, selectively targeting cytoskeleton components using drugs is rather challenging; instead, genetic modifications, e.g., gene knockdowns, can be employed [138]. Similarly, F-actin modulators can be targeted genetically to reveal their effects on cell mechanics, such as Rho GTPases Rho and Rac [89] or Ena/VASP [104]. In the latter two examples, a molecular basis for the cell mechanical changes coming with oncogenic transformation was explored.
Adherent cells can display highly varying stiffness values; these are further influenced by the substrate on which the cell is sitting on, with concomitant cell shape changes [48,70]. To control cell shape during cell mechanical testing, probed cells can be trapped within microwells [48]. In addition, 3D matrix stiffness [141,142] and compressive stresses arising from growth under confinement affect the mechanical properties of cells [135]. Since the mechanical properties of cells are less accessible within their 3D context, appropriate methods to assess them have to be chosen, e.g., passive or active microrheology or Brillouin microscopy [114,141,142]. Moreover, the presence of cell-cell contacts when cells are forming multicellular cell clusters can affect cell mechanical properties [16,135,143]. Resuspending adherently growing cells for mechanical probing has large effects on the cortical cytoskeleton [144]. On the other hand, it can be argued that the suspended state represents a “ground state” that is not affected by unnatural cell adaptions to stiff substates commonly used for 2D cell culture. Over the past years, also interesting links between cell mechanical properties and metabolism have become evident (reviewed in [145]. Since metabolic and mechanical changes are both hallmarks of cancer, more studies have to be conducted to reveal the underlying mechanistic links.
In sum, due to the substantial impact of above-discussed factors, i.e., cell state (cell cycle, metabolism, shape), context (2D/3D, matrix stiffness), cell preparation protocols (detachment of adherent cells, time in suspension) and measurement conditions (medium, temperature, deformation rates, force regime, cellular regions) on the mechanical phenotype of cells, the use of cancer cell mechanical markers in diagnostics requires reproducible and stringent protocols and thorough documentation of all experimental parameters.
2.6 Cancer cell responses to altered mechanical cues
As outlined in Section 2.1, the mechanical aberrations arising in tumors have multiple contributing factors, from increased ECM stiffness to cellular alterations that are likely to be sensed by neighboring cells. A vast number of studies have focused on stiffness sensing by cancer cells, which can also be studied experimentally through bioengineered 2D and 3D models. These studies have also been subject to recent reviews on the topic [34,146–148]. In the upcoming section, the focus will be on a less discussed subject, the influence of biofluid viscosity and shear stress on cancer cells.
The previous sections indicate that cancer cells exhibit softer mechanical properties compared to healthy cells. Notably, all the data in Figure 1 and Figure 2 were collected under static conditions, meaning no external flow was applied to the cellular environment, and conventional culture media were used. However, under physiological conditions, soft tissues — including tumors — are immersed in interstitial fluids, which can exert forces and stresses on surrounding cells. Interstitial fluids can locally have a viscosity 5 to 50 times that of cell culture medium due to the dissolution of extracellular macromolecules [24,149], making them even more viscous than lymph or blood (3–8 mPa · s) [150,151]. Interstitial fluids can also flow, imposing wall shear stresses on surrounding cells and tissues [24,25]. Although these effects have been known for years [24,152], they remain relatively understudied. Only recently, significant progress has been made in understanding how increased fluid viscosity influences cancer cell behavior.
2.6.1 Interstitial fluid viscosity
To increase the viscosity of the extracellular medium in 2D cultures, researchers used biopolymers like alginates, polysaccharides, mucins, and polyethylene glycols, adjusting their molecular weight and concentration [149,153–156]. These experiments aimed to replicate the properties of interstitial fluid in the tumor environment, achieving viscosities as high as 2 Pa s (2,000 times that of cell culture medium) [73]. Gonzalez-Molina et al. used viscosity-enhancing polymers to simulate extracellular fluids, conducting wound healing and cell spreading assays on liver cancer cells [153]. Surprisingly, despite the increased hydraulic resistance, cell motility was significantly enhanced on 2D substrates for both healthy and cancerous cells. Structural changes at the cell level, including cytoskeleton rearrangement, cytoplasmic expansion, and nuclear flattening were also observed [153]. Maity et al. later confirmed these findings, showing a 2.5-fold increase in migration speed compared to conventional cultures [154,157]. More recently, Bera et al. conducted an in-depth study, suggesting that the increased cell motility in high-viscosity environments is driven by a mechanotransduction pathway involving cytoskeleton-ion channel interactions, calcium influx, and enhanced cell contractility [149]. Importantly, Bera et al. demonstrated that this increased motility occurs in vivo using zebrafish, chick embryos and animal models, and affects metastasis. These findings suggest that higher local extracellular fluid viscosity could be a factor in cancer cell migration within the tumor microenvironment. Given the current knowledge of interstitial fluid viscosity, technologies for measuring their flow and stress in vivo for prognostic purposes are still unavailable to patients. However, developing such methods could offer valuable insights into key aspects of metastatic biology [34]. This contrasts with blood, which also plays a crucial role in the metastasis spread and has been extensively studied for its hemorheological changes related to cancer [25,158]. Well-established research shows that rheological alterations occur in advanced cancers and are often linked to disease stage and prognosis. For various cancers, including breast, lung, ovarian, and cervical cancers [158], it has been consistently observed that whole blood viscosity is higher in cancer patients compared to healthy controls [150]. This increase is attributed to elevated plasma viscosity and red blood cell aggregation, both of which are associated with metastasis development. A recent study on whole blood viscosity in patients with hepatocellular carcinoma found that both systolic and diastolic whole blood viscosities increased by 16% and 25%, respectively, compared to healthy individuals [150]. These increases are significant enough to improve clinical diagnosis for a range of cancers.
2.6.2 Interstitial fluid shear stress
Interstitial fluid flow in tumors is driven by pressure differences between blood and lymphatic systems. The flows of interstitial fluids and blood are governed by distinct physical models based on the medium they traverse. Interstitial fluid follows Darcy law for heterogeneous media, characterized by low velocities (1 μm s−1) and shear stresses in the range 0.01–0.1 Pa. Blood, in contrast, follows a pulsatile Poiseuille flow with velocities 10 to 105 times higher than interstitial flow, with shear stresses range from 0.1 to 10 Pa [25]. As a result, the behavior of cancer cells within a tumor, or circulating in the bloodstream varies significantly [24,34]. Recent studies have shown that cancer cells can respond to a wide range of interstitial stresses [159–163], with experiments conducted using in vitro microfluidic devices designed to replicate the tumor microenvironment. Most commonly, single microfluidic channels are used, where cells line the channel walls and a controlled flow is applied. Relevant work includes that of Calibasi Kocal et al., who developed a microfluidic platform capable of generating low shear stress around 10–⁵ Pa on esophageal cancer cells, showing that laminar flow induces epithelial/mesenchymal hybrid transition and increases mechanotransduction protein expression, in contrast to static cultures [159]. Other recent reports explored how shear stress in the range of 0.005 Pa to 0.5 Pa influences ovarian, breast, and prostate cancer cell behavior through the activation of mechanosensitive sodium and calcium ion channels, enhancing motility and tumor growth, or allowing tumor cells to evade high shear regions during intravasation [162,163]. The impact of fluid shear stress on cervical cancer cells also revealed that moderate shear stress boosts cell proliferation and resistance to chemotherapy, while high shear stress suppresses growth, indicating that fluid shear stress significantly influences cancer cell behavior and drug resistance during metastasis [160].
3. Electrical Properties of Cancer Cells
The investigation of electrical properties of cancer cells is a rapidly evolving area with promising implications for cancer diagnosis, prognosis, and monitoring of cancer treatment [164–166]. Compared to normal cells, cancer cells exhibit distinct morphological and functional features [167–170] due to the accumulation of charged metabolites and ions, ion channel activity, and alterations in membrane composition and cell size [171–174]. These biomolecular variations can serve as versatile markers for cell proliferation, apoptosis, adhesion, cell cycle determination, and other cellular processes but also substantially affect the electrical properties of cells.
One of the hallmarks of tumor metabolism, known as the Warburg effect, has a high influence on the pH values of tumor tissues and cell cytosol due to the activation of the ion channel proteins and accumulation of the extracellular lactate. For instance, the pH changes and activity of ion channels lead to the depolarizing of the cell membrane potential [175,176]. Thus, deregulation of the ion channels and transporters results in uncontrolled tumor cell growth and spread [177].
As it is difficult to attribute the changes on a molecular level with the resulting electrical signal, the electric properties of normal and tumor cells are often correlated to key biological functions or cell phenotypes. In this way, the influence of the cytoskeleton on the resistance of the inner part of the cell [178] has been investigated. Also, the observation throughout the cell cycle revealed differences in phases, which have been attributed to the increase of membrane surface area and the rearrangements of large structures inside the cell [179]. Interestingly, there are assumptions that morphological changes can be associated with the electrical properties of the cells [180]. In particular, enhanced plasma membrane damage has been associated with increased motility and invasiveness of tumor cells [181], and the electrical measurement of the damaged membranes could be used as a marker of tumor aggressiveness. Furthermore, morphological features such as cell size, and cell functional properties, e.g., necrosis and apoptosis, can also be detected at low frequencies [182].
Understanding the ‘electrical signatures’ of cancer provides valuable insights into cellular physiology and transformation, tumor heterogeneity, and proliferative behavior [165,183]. Tracking electrical parameters, such as conductivity and permittivity, enables diagnosis, monitoring, and treatment surveillance by identifying different cell types and distinguishing between healthy cells and those exhibiting abnormal behavior. Therefore, it is crucial to explore how the changes in electrical properties are related to cancer types and cancer development.
3.1 Biological basis of the tumor-specific cell electrical properties
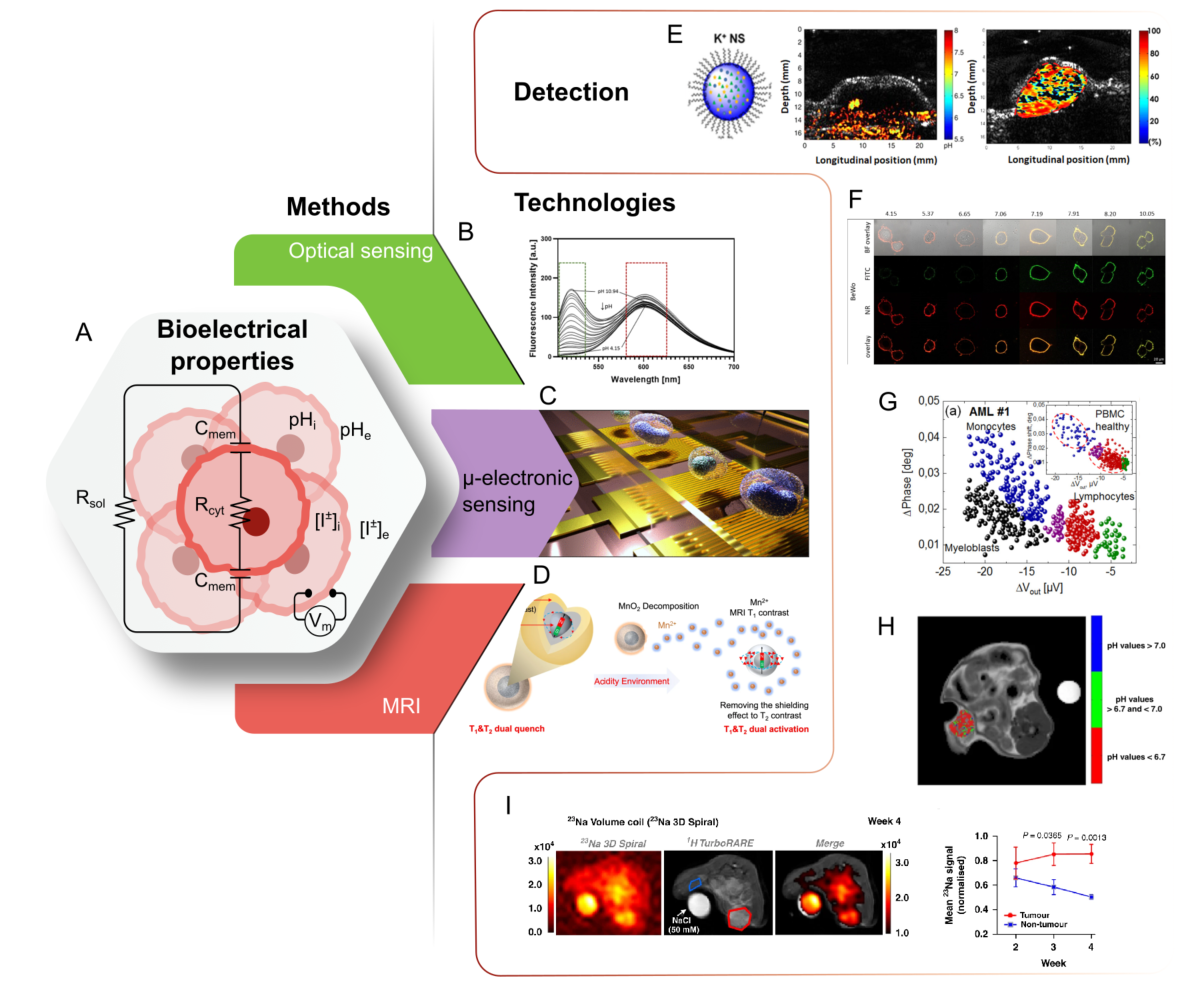
The electrical properties of cells reflect their physiology and pathology, influenced by both their intracellular and extracellular environments [183]. When applied to cancer, tumor-specific cell electrical profiles emerge, differing significantly from those of healthy cells. Particular interest represents changes in the mechanisms regulating pH, ion levels, and membrane potential, leading to the alteration of various cellular processes and abnormal behavior (Figure 3).
3.1.1 pH
Dysregulation of pH balance, intracellularly and in the tumor microenvironment, contributes to uncontrolled proliferation and immune evasion [183,184]. Variations in pH values become fundamental to the survival of cancer cells. Note, that the intracellular pH in healthy cells (6.99–7.05) is kept lower than the extracellular pH (7.35–7.45), while cancer cells display a “proton gradient reversal” with extracellular values (6.2–6.8) more acidic than the intracellular pH (7.2–7.8) [185]. Unlike normal cells, tumor cells thrive in environments with high lactate and hydrogen ions (H+) production, conditions that would typically induce cell death [186]. However, tumor cells counteract this threat with the enhanced expression of several intracellular pH-regulating systems [187]. In contrast to tumors, healthy cells can barely survive in this hostile environment [186]. pH changes in cancer cells influence the activity of cellular channels, leading to different ionic concentrations and depolarizing the cell membrane potential [175]. Additionally, studies have shown a relationship between intracellular pH changes and the dynamics of microtubules and actin filaments, further highlighting the impact of pH on cancer cell function [188,189].
3.1.2 Role of dysregulation of ion channels
Dysregulation of the functions of ion channels and transporters is a condition that supports the uncontrolled growth and spread of cancer [177]. The role of ion channel malfunction in cancer progression and metastasis is currently of great interest, considering that not all tumors share the same pattern of ion channel expression [183]. Overall, changes in calcium, sodium, and potassium promote a cascade of events that result in uncontrolled proliferation, abnormal cell physiology, and ultimately contribute to the formation of aggressive, metastatic tumors [183].
Precise concentration and localization of calcium (Ca2+) ions govern cell proliferation and apoptosis [196]. Cancer cells capitalize and maintain calcium levels that avoid death. Calcium levels in tumors also favor cell proliferation and angiogenesis, which enhances nutrient distribution and growth [196–198]. Cancer and wounded cells are known for excessive intake of water and sodium (Na+) [199,200]. This feature affects the cell towards a bigger size and more spherical shape. This altered shape influences cell signaling contributing to cancer progression and metastasis [200]. Cancer cells often exhibit enhanced expression of potassium (K+)-calcium (Ca2+) channels [183]. Altering the cell's response to hypoxia and modifying cell adhesion, migration, and apoptosis, tumor progression is favored [201–203]. The aforementioned ionic changes also influence the membrane potential. Cancer cells have a more depolarized resting membrane potential (Vm) compared to healthy cells, contributing to an increased proliferation and migration capacity of tumor cells [204].
3.1.3 Membrane potential
The membrane potential, determined by the ion concentrations, is highly affected by ion channels' permeability, expression, and activity. Cancerous cells show a tendency to have a more depolarized Vm compared to healthy cells. This depolarization is associated with poorer patient prognosis. Depolarized cells increase their proliferative behavior in cancer, influencing the cell cycle progression [204,205]. Hyperpolarization is required to initiate apoptosis, and the depolarized state disrupts this process increasing the survival rate of cancer cells [204,206]. Furthermore, the depolarized phenotype of cancer cells is associated with stem-cell-like behavior characterized by self-renewal, inhibition of cell differentiation, and migration. All these factors contribute to increased tumor aggression and metastasis [204].
3.2 Impedance of cancer cells
The transformed state of cancer cells is also reflected in a change in membrane capacitance and conductivity (Figure 3A). Cancer cells frequently display an abundance of sialic acid-rich glycoproteins, resulting in a more negative surface charge [200,207,208]. This acts as an electrical shield that protects cancer cells from negatively charged immune effector cells [200]. The changes in composition, shape, and permeability anticipate a differential charge storage capacity with respect to healthy cells [209]. This relative difference is a more reliable marker for malignancy than the absolute capacitance of the cells [209]. In support of this conclusion, studies have linked variations in membrane capacitance to patient survival rates in head and neck cancer [210]. Beyond capacitance, it is observed that conductivity also has increased values in cancer development and malignancy; this suggests that impedimetric measurements can be used to assess the prognosis of cancer based on impedimetric biomarkers [165,211].
Impedance (Z) is measured by applying an alternating current (AC) potential enabling to distinguish cancer and non-cancerous cells in a real-time and label-free manner. To understand the concept, a rather simplified explanation will be used; for a more comprehensive understanding, please consult Lazanas et al. (2023) [212]. Impedance is a complex property of materials and can be separated into a real (resistance –R) and imaginary (reactance –X) part; with its complex character and frequency dependence, it provides the possibility to derive multiple other cell-specific parameters that can be used to gain important information about cell processes and properties.
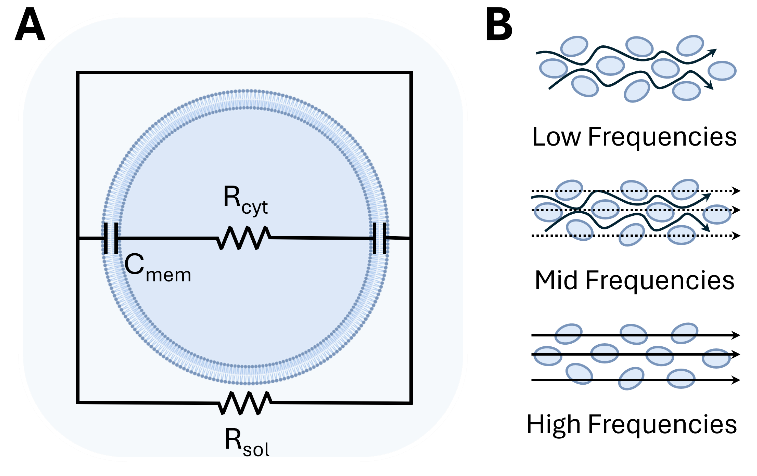
In common impedance measurements, the magnitude (|Z|) and the phase angle between voltage and current (θ) are determined, which can be used to derive the capacitance (C), the ability of a system to store electrical charge, conductivity (σ), the ability of a cell to conduct electric current, and permittivity (ε) the ability to polarize in response to an electric field (Table S1). All these properties have a high potential to be used to improve the understanding of cellular processes. The simplified models are used for the measurement evaluation, and the equivalent circuit model (Figure 4A) is one of the most common. The cell compartments are simplified as elements in an electrical circuit, with properties that have minimal impact on the overall signal being neglected. This model consists of two main components: the cell membrane, which primarily contributes capacitance due to its bilayer structure and is represented as a capacitor (Cmem), and the cytosolic side, which exhibits resistive behavior and is modeled as a resistor (Rcyt). Both elements are arranged in parallel and combined with components that represent the surrounding environment, such as the ECM and medium [213].
The frequency plays a key role in understanding various properties, as it primarily influences the interaction of current [214]. Parts of the cell affected by current can be divided into three frequency ranges: low frequencies, where the cell acts as an insulator with no current transmission but allows cell size determination; mid frequencies, where partial interaction with the cytosolic side occurs, reflecting membrane capacitance; and high frequencies, which penetrate the cell and interact most with the cytoplasm (Figure 4B) [214,215].
3.3 Methodology
3.3.1 Optical sensing techniques
Fluorescence microscopy and spectroscopy are well-developed and state-of-the-art techniques for imaging the inner cell processes at high resolution. These techniques, relying on light absorption and emission at a larger wavelength, are carefully adapted to interact with specific cell components [216]. By analyzing light intensity, wavelength, and distribution, it is possible to extract information about concentration, binding state, and environmental conditions experienced by the probe in the tumor or cancer cell (Figure 3B) [217,218].
Fluorescent pH indicators, such as SNARF-1 and the genetically encoded pH sensor SypHer2, have been used to study the reversed pH gradient in cancer and assess the efficacy of pH-regulating therapies [186,219]. By calculating the fluorescence ratio at two different wavelengths of the ratiometric dye SNARF-1, the extracellular pH of a tumor was measured in vivo [186]. In addition, genetically encoded pH nanosensors can be designed to trace single extra- to intracellular gradients and monitor intracellular pH response to different stimuli, such as treatment with pharmaceuticals. One recent example represents the nanosensor consisting of biocompatible polystyrene nanoparticles loaded with the pH-inert reference dye Nile red, and is surface-functionalized with a pH-responsive fluorescein dye. The nanosensor is equipped with a targeting moiety and can adhere to cell membranes, allowing direct measurement of extracellular pH at the cell surface (Figure 3F) [190].
Ion dynamics can also be studied through fluorescence methods. Ion concentration transients and waves have been detected through calcium-related markers probing the long-distance communication between cancer cells in human colon and prostate cancer cells [218]. Further efforts to quantitatively evaluate the tumor microenvironment concentration of potassium ions in vivo have been realized through the combination of fluorescent nanosensors and photoacoustic emission (Figure 3E) [192,193]. This analysis revealed significantly higher levels of potassium ions (29 mM) in the microenvironment compared to the surrounding muscle tissue (19 mM).
By tracking the expression of K+ channels tagged with green fluorescent protein (GFP) probes, researchers have studied the effects of manipulating the membrane potential in cancer cells. It was found that hyperpolarization of the membrane potential through K+ channels overexpression increases breast cancer cell migration, invasion, tumor growth, and metastasis [202]. The increased polarization creates conditions to upregulate cadherin-11, leading to metastatic behavior [202].
3.3.2 Magnetic resonance imaging
The electrical properties of cancer cells have been studied through the magnetic resonance imaging (MRI) technique that utilizes the interaction in high-intensity magnetic fields of magnetic moments of atoms and radiofrequency pulses [220]. It can utilize contrast agents with optimized nuclei relaxation times that enhance the contrast between specific ions, molecules, and targets (Figure 3D) [191,221].
While traditional imaging often overlooks ions, researchers have recognized that altered ion levels within tumors hold valuable diagnostic and prognostic information. Previous investigations have hinted at a prevalence of higher concentrations of sodium ions Na+ in cancer cells compared to the surrounding healthy tissue [199]. After analyzing orthotopic breast cancer regions with MRI combined with diffusion-weighted imaging (DWI), it was possible to confirm this observation and identify that the excess of Na+ is concentrated intracellularly in cancer cells (Figure 3I) [195]. By employing both techniques, enhanced levels of sodium ions were linked to increased cellularity in breast cancer models that serve as a promising biophysical marker in early tumor development [195].
Advanced MRI-based chemical exchange saturation transfer (CEST) has been shown as a promising tool for the diagnosis of metastases potential in primary tumors based on spatial acidosis studies (Figure 3H) [194]. A pH-responsive dual-mode MRI contrast agent was also demonstrated to be effective in detecting metastasis in liver tumors with sizes of 0.5 mm when using the T1-T2 dual-modal MRI contrast agent for cancer imaging [191].
3.3.3 Microelectronic probe
While previous methods can infer cellular electrical properties, using microelectronic techniques provides a direct characterization of these parameters (Figure 3C). Direct electrical measurements offer the advantage of real-time quantifiable dynamics of localized events [222]. Based on the fact that each biological system has distinct dielectric properties, it is possible to use them as relevant quantitative biophysical markers in cancer prognosis [223].
Microelectrodes have been engineered in recessed-tip, and double-barreled configurations to facilitate negligible perturbations during sensing with small tip sizes and simultaneous measurements of the membrane potential and pH, respectively [224,225]. Due to the compatibility with microfabrication techniques, it has been shown that it is possible to fabricate arrays of sensors for high throughput tracking of the heterogeneity of pH changes in cancer cells [226]. The activity of voltage-gated ion channels has also been related to metastatic potential in prostate cancer cells through the patch clamp technique [227]. The direct influence of bias voltage and sensing through integrated sources makes possible the control and regulation of specific ion channels [228].
3.3.3.1 Electrical impedance spectroscopy (EIS)
The use of electrical impedance spectroscopy (EIS) for analyzing biological samples has a high potential due to its sensitivity and specificity. A variety of electrode designs have been developed to optimize the precision of measurements [229]. Frequency sweeping enables the detection of changes in response across a broad range of AC frequencies, providing a comprehensive view of how the system behaves at different frequencies. Depending on the necessary dynamics that must be detected, the range of frequencies can be chosen, and the number of frequencies has to be adjusted accordingly in a relatively quick manner. A clear limitation is the time resolution and capability of complex measurements of the used device.
EIS has already shown its versatile character in cancer research, as already mentioned [229]. Multiple studies have shown that EIS can differentiate between cancerous and benign cells [167,230,231]. When treated with anti-cytoskeletal drugs, breast cancer cell lines with increasing malignancy displayed distinct resistive behavior, highlighting the impact of microtubules and actin on the cell impedance [232]. Cell behaviors, such as migration and invasion, can be monitored, with migration detected by an increase in impedance as cells move onto the electrodes [233,234], and signal changes across multiple frequencies can be used to quantify invasion. As cancer cells degrade the EMC, their higher impedance compared to the ECM was interpreted as evidence of invasion [235].
3.3.3.2 Electrical impedance cytometry (EIC)
The ability to polarize and measure the conductivity of cells and their components can be assessed through electrical impedance cytometry. This technique, based on the excitation of an electric field (in the kHz to MHz range) in the area between micro- and nanoelectrodes, can differentiate the capacitive and conductive properties of healthy and cancer cells [223,236]. This approach has proved the successful distinction between healthy tissue and breast carcinoma tissue as a prospective early diagnostic tool [165]. This technique can be extended to in vivo tissue, using an array of 90 electrodes to map the dielectric properties directly in patients; researchers performed a direct tomography extracting the cues of abnormal tissue [237].
Electrical impedance cytometry (EIC) offers a complementary approach to EIS for analyzing the electrical properties of cancer cells. Unlike impedance spectroscopy, which typically measures the impedance of monolayers or tissues over a range of frequencies, impedance cytometry focuses on individual cells and single frequencies in real-time. This technique utilizes microelectrode arrays to measure changes in electrical impedance as cells pass through a microfluidic channel [238]. The advances in device integration have allowed the development of electrical analogous to fluorescence cytometry assays. Impedance cytometry uses capacitive-like electrodes integrated into microfluidic channels that analyze the heterogeneity of single cells through their impedance signatures [238]. This label-free method has been demonstrated in a pilot study to identify cancer cells in the peripheral blood of patients with acute myeloid leukemia (Figure 3G) [164]. Another pilot study demonstrated that the differentiation of cancer cells in human peripheral blood in patients with acute myeloid leukemia was possible [164]. It was also demonstrated that bladder cancer cells encapsulated in microdroplets generated by a microfluidic device were successfully discriminated [239].
Another property that is often determined in EIC is opacity, which represents the transmission of electromagnetic fields (electrical penetration) and can be used to study the cell membrane and size, by plotting the absolute values of the impedance at low and high frequencies against each other [64,240]. These techniques offer a high potential for creating high-throughput miniaturized devices that could be used in point-of-care settings. EIC is particularly advantageous for drug and marker screening. Machine learning is often used to automate the EIC data analysis process and avoid human bias [238,241].
3.3.3.3 Manipulation of cells with dielectrophoresis (DEP)
Dielectrophoresis (DEP) is a technique to manipulate cells based on their dielectric properties when exposed to a non-uniform electric field. Single cells can be affected by negative DEP which deflects them from areas with a high electric field, while positive DEP attracts them towards high-field regions [242].
This method enables the separation and analysis of cancerous cells from healthy ones due to their distinct electrical characteristics [243]. DEP can be utilized for the isolation of circulating tumor cells (CTCs) from blood [244], offering a label-free technique for liquid biopsy analysis.
4. Thermal Properties of Cancer Cells
4.1 Theoretical background and determination of the basic parameters
The thermal properties of healthy tissue cells and tumor cells can differ. These differences arise due to the distinct metabolic activities, structural composition, and microenvironment of the two cell types [245]. Tumor cells often have higher metabolic rates than healthy cells [4,246]. This increased metabolic activity can lead to excessive heat production in tumor tissues [247]. This phenomenon is partially due to the Warburg effect, where cancer cells preferentially use glycolysis over oxidative phosphorylation, even in the presence of oxygen [247]. In addition to the increased metabolic activity, the rapid proliferation contributes to the elevated heat generation, the so-called thermogenesis [248]. Tumor tissues may have different compositions compared to healthy tissues, including variations in cell density, ECM, and vascularization [249]. These differences can affect the thermal conductivity κ of the tissue [250]:
where κ is the thermal conductivity of the tissue []; Q is the rate of energy transfer (or heat flow) through the tissue [W]; A is the area of the surface through which heat is transferred [m2]; ΔT is the difference in temperature across the tissue [K]; L is the thickness of the tissue sample or the distance over which the temperature difference is measured [m].
To access the structural differences in cells, including variations in cell density and the organization of cells and extracellular matrix, one can define the thermal diffusivity α [250]:
where α is the thermal diffusivity of the bulk cell culture []; κ is the thermal conductivity of the bulk cell culture ; ρ is the density of the tissue or cell culture []; cp is the specific heat capacity of the tissue [].
Tumor tissues might have a lower thermal diffusivity compared to healthy tissues because of their denser and unorganized structure. The specific heat capacity of a tissue depends on its biochemical composition, including the amounts of water, lipids, and proteins [251]. Changes in the biochemical composition of tumor cells, such as altered levels of these components, can affect their specific heat capacity. Consequently, thermal diffusivity measurements can provide insights into the structural and compositional differences between healthy and cancerous tissues. Tumor cells may have altered biochemical compositions, affecting their specific heat capacity [251].
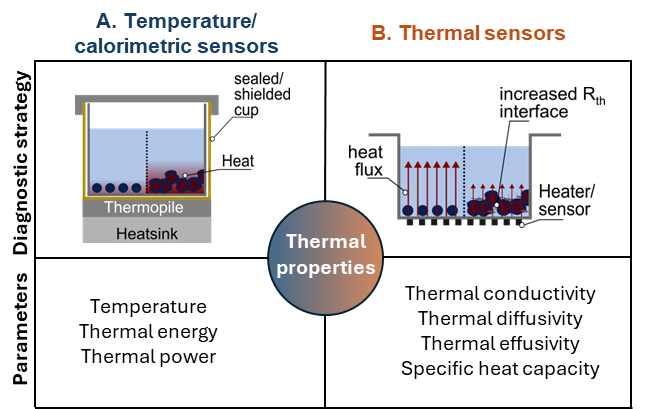
There are two main groups of thermal sensors to detect these physiological differences, temperature/ calorimetric sensors, which measure heat generation or heat transfer, see Figure 5A), thermal sensors, which measure physical properties such as thermal diffusivity, thermal effusivity or thermal conductivity, see Figure 5B). Indirect thermal sensors that derive signals such as force pressure, flow rate, or acceleration do not yet play a major role in cancer research.
While the potential of these thermal properties as prognostic biomarkers is extensive, research in this field has not garnered significant attention. However, some promising techniques have been explored, particularly those utilizing calorimetry and heat transfer methods. It is important to note that the application of thermal sensors has so far been mainly limited to in-vitro studies under controlled laboratory conditions and has not yet been widely implemented in in-vivo or clinical settings. The following sections discuss these methodologies and introduce a recent strategy with potential applications in diagnostic settings.
4.2 Methodology
4.2.1 Calorimetry
Calorimetry is a label-free technique used to measure the heat generated or absorbed by biological samples, providing insights into their metabolic activity [252]. The heat generated can reflect physiological processes in normal cell cultures but can also be employed to detect cellular responses to various stimuli, such as drugs, environmental changes, or stressors, offering a versatile tool for studying cellular behavior in different conditions. Growth of hepatocarcinoma tumor microtissues (based on HepG2 cell line) releasing a maximum thermal power was monitored, which corresponds to a diameter increase of the microtissues from ca. 100 to 428 μm [252]. Isothermal microcalorimetry, in particular, was also used to investigate the influence of enriched media compared to normal medium on neuroblastoma cells [253]. The spiking of fructose, glucose or fructose/glucose to the medium led to increases in the metabolic activity of the cells [253]. These studies also show that tested glioblastoma cells (Kelly and SH-EP Tet-21/N) preferred fructose metabolism over glucose metabolism, a beneficial adaption of these cancer cells under low oxygen and low nutrient conditions [253]. While differential scanning calorimetry (DSC) is known as a sensitive technique in the field of phase transition studies, during the last decades, it has also been recognized as a tool for cancer diagnosis and monitoring [254–256]. DSC has been tested as a tool for lung cancer patient diagnosis and prediction in clinical settings [257]. Different lung cancer subtypes and stages were successfully distinguished from control patients [257]. With further optimization DSC could provide an accurate, non-invasive, and radiation-free strategy for state-of-the-art low-dose CT scans [257].
4.2.2 Heat transfer method (HTM)
HTM is a thermal transducer platform that monitors the thermal interface resistance Rth between the solid chip and the supernatant liquid [258]. When combined with surface-imprinted polymers (SIPs), HTM has been used to distinguish between human breast cancer cell lines MCF-7 and leukemia Jurkat cells [259], and to detect differences in their glycosylation patterns using modified Chinese hamster ovarian cells [260]. The model allowed excessive expression of the transmembrane protein mucin-1 (MUC1) and control of its glycosylation by varying culturing protocols [260]. The selective differentiation between cells expressing the MUC1 protein and cells that do not express MUC1 and MUC1 glycosylated and MUC1 non-glycosylation indicate significant differences in the imprints [260]. HTM has also been effective in monitoring the quality of cell cultures over time, as demonstrated by comparisons between adherent breast cancer cell culture (ZR-75-1a) and suspension culture (ZR-75-1s) [261].
4.2.3 Modified transient plane source (mTPS) sensor
Another noteworthy technique is the mTPS sensor systems, which can combine aspects of calorimetry with the monitoring of changes in thermal properties at the chip interface. Although not yet applied to cancer detection, mTPS has been used for real-time quantification of yeast cell numbers and the metabolic activity of cultures [262]. The label-free and indirect sensing strategy is suited for bi-modal sensing, revealing both thermal and electrical information about the sample [263,264]. The integration of mTPS into microplates for high-throughput screening or microfluidic chips has also been explored [265], and therefore holds great potential for future life science applications.
5. Tumor Heterogeneity
5.1 Intertumoral and intratumoral heterogeneity and cancer stem cells (CSCs)
Tumors are complex living systems where tumor cells and their microenvironment are regulated in a bilateral feedback loop manner. As each patient is unique, so are their tumors, which possess intertumoral (between tumors) heterogeneity [266,267]. Molecular fingerprinting of individual tumors by protein analysis, mRNA expression, or DNA sequencing serves as a basis for the development of cancer biomarkers - measurable indicators of tumor presence and progression [268]. In addition to the difference between the patients, each individual tumor possesses a certain level of intratumoral (within a particular tumor) heterogeneity attributed to the tumor cell phenotypes as well as genetic and epigenetic traits [269]. Intratumoral genetic diversity is a result of genomic instability, which is an increased frequency of mutations during cell division. Genomic instability is attributed to many types of tumors [270]. In tumor bulk, most cancer cells have limited proliferative potential. Thus, not all acquired mutations are passed across multiple cell generations and fuel tumor evolution. In contrast, only a subpopulation of tumor cells called CSCs possesses unlimited self-renewal potential and the ability to differentiate into other tumor and non-tumor cell subsets [271]. Due to their unlimited self-renewal potential and ability to recapitulate all tumor cell populations, these cells maintain tumor growth and, therefore, are considered a unit of tumor evolution. A general consensus is that tumor cells with CSC properties not only maintain the primary tumor growth, but evolve, disseminate, and give rise to tumor metastasis [272]. The populations of CSCs can be identified and isolated using different plasma membrane and intracellular protein molecules serving as CSC markers, such as CD44, CD133, integrin α2β1, and others [273–275]. CSC-related biological markers are the focus of many clinical trials as promising prognostic indicators and therapeutic targets [276–279]. Furthermore, CSCs can be differentiated from non-CSCs by physical properties such as their size, viscoelasticity, and electrical properties [165,280–283].
CSCs produce cell clones that expand during tumor development and possess different genetic, epigenetic, and functional features [271,284]. Tumor heterogeneity depends on the number of CSCs that contribute to tumor growth. At the same time, genomic instability, the number of acquired genomic alterations, and the heterogeneity of CSCs are increasing during tumor progression. The evolutionary dynamics and spatial tumor heterogeneity can be analyzed by different approaches, such as longitudinal sampling, spatial tumor biopsies, and multi-omics profiling, including whole genome sequencing (WGS), whole exome sequencing (WES), RNA sequencing (RNAseq), assay for transposase-accessible chromatin sequencing (ATAC-seq), single-cell RNA sequencing (scRNAseq) and single-cell mass spectrometry, spatial proteomics and transcriptomics, etc. [285,286]. In addition to their distinct molecular and functional characteristics, different clones composing a given tumor possess specific functional features and physical parameters such as cell size, viscoelasticity, and cytoplasmic viscosity, and have different impacts on the microenvironmental components, including hypoxia, acidity, extracellular fluid viscosity and matrix stiffness [11,287].
5.2 An interplay between biological and mechanical heterogeneity and tumor stemness
5.2.1 The mechanotransduction signaling
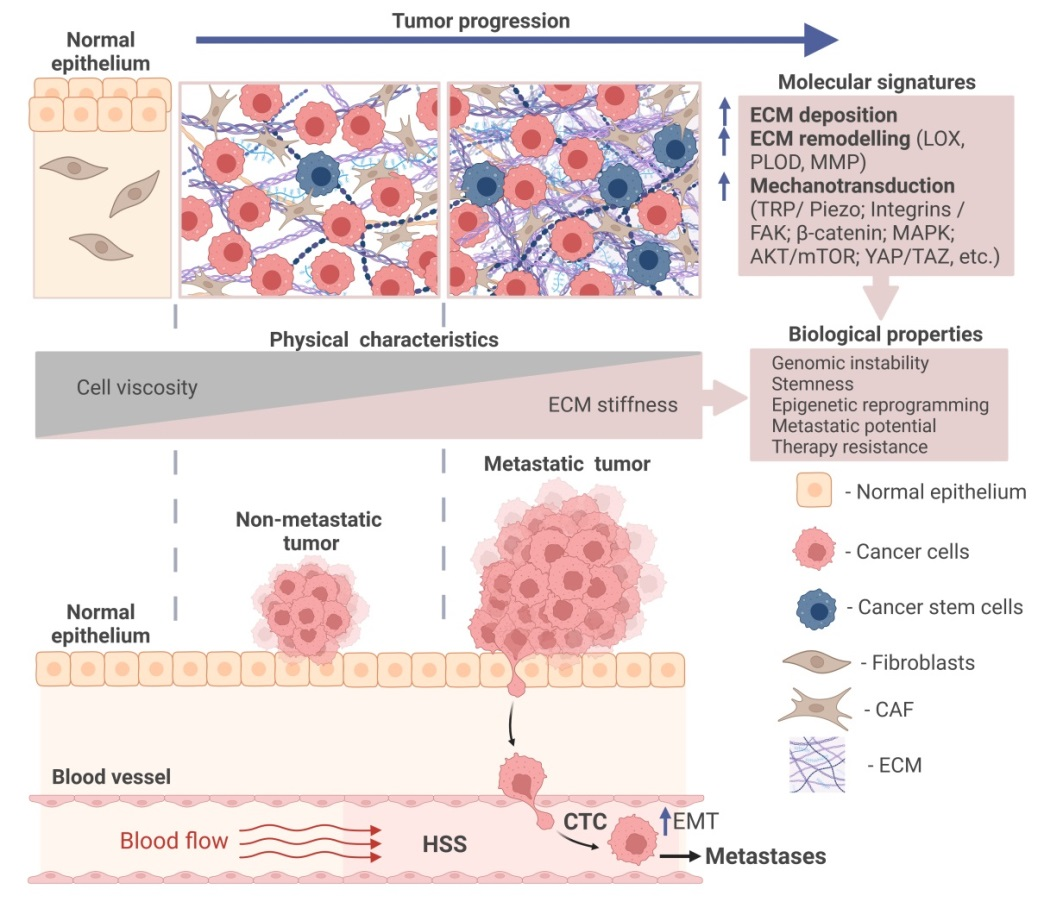
Tumor growth is accompanied by biomechanical changes in the tumor microenvironment, including an increase in matrix stiffness generated by ECM components, and the accumulation of solid stresses from compression or tension during tumor growth [288]. Analysis of the genomic mutations in cancer of different types suggested that tumor tissue stiffness correlates with the scale of genomic variations and intratumoral heterogeneity [289]. This increased genomic variation can be associated with increased DNA damage in response to the cell and nuclear squeezing upon cell proliferation in a stiff microenvironment or migration through tiny pores of the collagen-enriched ECM or basement membranes [289,290]. The stiffness of tumor tissues depends on the deposition of the ECM components such as collagen, laminin, elastin, fibronectin, and glycosaminoglycans (GAG) by cancer-associated fibroblasts (CAFs) and tumor cells and ECM remodeling by enzymes such as matrix metalloproteinases (MMP), procollagen-lysine, 2-oxoglutarate 5-dioxygenase (PLOD) and lysyl oxidase (LOX) secreted by tumor cells and tumor-associated stroma cells [22,291]. Mechanical stress induces actin stress fiber reorganization in the tumor cells, consequently affecting cell elasticity [292,293]. Mechanical stress also induces large-scale chromatin remodeling and changes in gene expression profiles [294]. Mechanical stimuli can activate cell membrane mechanosensors such as transient receptor potential (TRP) channels and Piezo ion channels [295], integrin receptors [296], G-protein coupled receptors (GPCR) [297] and cadherins [296,298], and corresponding cytoplasmic mechanotransducers including RhoA-mediated actin dynamics [299], focal adhesion kinase (FAK) complexes, transcription factors (e.g., yes-associated protein (YAP)/WW domain-containing transcription regulator 1 (TAZ), mechanistic target of rapamycin complex 1 (mTORC1) [300], phosphoinositide 3-kinase (PI3K)/Akt [301], and β-catenin [298], which induce other downstream signaling mechanisms in tumor and stroma cells [302,303]. The mechanotransduction signaling is activated in most types of cancer. It is involved in the regulation of different aspects of cancer progression, including tumor growth, therapy response, immune evasion, metastasis, and stemness [304].
5.2.2 Mechanical properties of CSCs
Indeed, for many cancer types, CSCs are shown to be the dynamic and plastic cell populations. The epigenetic changes, newly acquired mutations, and microenvironmental factors such as hypoxia, nutrient availability, physical parameters including stiffness, shear stress, and microenvironment architecture, as well as different types of treatments, can induce a bi-directional transition between CSC and non-CSC states. CSC surface markers such as CD44, CD133, integrin α2β1, and integrin β4 directly interact with collagen and GAG hyaluronan (HA) and induce intracellular signaling critical for CSC maintenance such as FAK, Akt/mTOR, MAPKs and β-catenin [305–308]. Indeed, increasing stiffness leads to the enrichment of CSC populations in breast [281], liver [309], and lung tumors [282], as reviewed recently [310]. High stiffness triggers TAZ-mediated mechanotransduction [281] and activates the YAP/β-catenin-dependent transcriptional program and expression of the reprogramming factors Nanog, Oct4, and Sox2 [281,282]. Integrin-mediated activation of FAK and downstream pathways, including AKT/mTOR signaling, contribute to CSC regulation through matrix stiffness [311]. Mechanotransduction mediated by Piezo ion channels is critical for the dissemination of cancer cells [162,312–315], and the high expression of the Piezo mechanoreceptors was correlated with worse clinical outcomes and metastases in many cancer types [313,315–319]. Piezo proteins, the mechanosensitive ion channel, are sensing cell deformation and transforming mechanical stimuli such as pressure or shear stress into biochemical processes. In particular, activation of Piezo channels leads to the influx of extracellular ions, mainly Ca2+, triggers the RhoA pathway regulating actin cytoskeleton [299,320], and activates different intracellular signaling mechanisms, including integrin/FAK, Akt/mTOR, and MAPKs [321–323]. Piezo proteins regulate the maintenance of CSC populations. Genetic deletion of Piezo1 inhibits CSCs in glioblastoma and colon cancer preclinical models [317,324]. The expression levels of Piezo1 or Piezo2 correlate with CSC-related protein markers and transcriptional signatures in patients with colorectal and gastric cancer, respectively [318,324]. TRP family of mechanoreceptors also plays a critical role in the regulation of CSCs, as recently reviewed [325].
Given the importance of mechanotransduction for CSC maintenance, it is unsurprising that CSCs can be identified and isolated based on their intrinsic mechanical properties: several studies demonstrated that CSCs are a mechanically soft population compared to non-CSC counterparts. The soft tumor populations were isolated from breast cancer cell lines by using microfluidic devices and characterized using in vivo models. These experiments demonstrated that soft tumor cells are more metastatic and tumorigenic in vivo and possess CSC properties [326]. The previous studies reached a consensus conclusion that metastatic tumor cells are much softer than non-metastatic cells [11,47,65,106], and as discussed in Section 2. Tumor cell softness was associated with an oxygen deficiency called hypoxia, another feature of aggressive, metastatic, and therapy-resistant tumors [11,327]. In each individual tumor, different spatially defined areas may be perfused differently by functional blood vessels [328]. Therefore, hypoxia has substantial inter- and intratumoral heterogeneity [329]. The hypoxic microenvironment promotes CSCs and metastatic properties, and one of the key mechanisms of this regulation is the activation of the transcriptional program driven by hypoxia-inducible factors (HIF). Both hypoxia and ECM stiffness induce CSCs, tumor growth and metastases through the mechanotransduction pathways such as β1-integrin/integrin-linked kinase (ILK)/PI3K/Akt mechanism [330]. In a feedback loop of this interplay, Piezo1-mediated mechanotransduction is shown to induce HIF1α expression [313].
5.2.3 Dynamic changes of the CSC mechanical properties in the metastatic cascade
A subset of CSCs determines metastatic growth through sequential steps, including tumor cell intravasation into the bloodstream, dissemination of the CTCs to distant organs, extravasation at the distant site, and initiating metastatic tumor development [246,331]. At the initial stage of the metastatic progression, immobile cancer epithelial cells acquire mesenchymal phenotype and the ability to migrate in a process called epithelial-mesenchymal transition (EMT) [332,333]. The EMT process is associated with generating a range of intermediate cell phenotypes, including both epithelial and mesenchymal features. EMT is positively associated with other tumor hallmarks, such as genomic instability and hypoxia. This developmental mechanism is associated with an epigenetic reprogramming of the bulk tumor cells into CSCs [333,334] and contributes to the heterogeneity of CTCs [335]. Therefore, EMT is considered to be a key driver of intratumoral heterogeneity [336]. The ECM stiffness activates the EMT program via Piezo1/2-mediated Ca2+ influx and mechanosensitive ephrin type A receptor 2 (Epha2)/Lyn protein complex followed by the activation of the TGF-β and Akt signaling pathways and transcriptional program driven by Twist1 and Snail [214,316,337–340]. Consequently, the activation of the EMT program softens the cytoplasm [341,342], induces actin reorganization, and increases actomyosin traction forces [342], playing a key role in cell migration [343], and, therefore, facilitates tumor cell invasiveness. At the intravasation stage of metastatic progression, the highly migratory and invasive tumor cells leave the primary site and invade through the basement membranes to the blood of lymphatic vessels. The stiffness of the ECM is a limitation encountered by invading cells, and only a subpopulation of tumor cells resistant to deformation and able to move through stiff microenvironment and dense basement membrane might survive and propagate metastases. As discussed in Section 2, highly metastatic cells have lower elasticity and viscosity compared to less metastatic tumor cells. The EMT program induced by stiff microenvironments makes prostate tumor cells more migratory than in soft substrate, enabling them to escape. The physical cell compression upon squeezing through narrow gaps in the wall of blood vessels is associated with nuclear envelop rupture (NER), temporary disruption of a lipid bilayer surrounding cell nuclei [290]. NER is associated with DNA damage and is potentially leading to the genomic heterogeneity of the metastatic cells [344]. The recent studies described the adaptive longitudinal dynamics of single tumor cell viscoelasticity under different pressures in the microvessels and softening of tumor cell nuclei during transendothelial migration [345,346]. The cells with softer nuclei have advantages in cell migration but a higher possibility of DNA damage and genome instability [347]. In addition to its effect on DNA integrity, the stiff microenvironment also induces mechanical adaptations of tumor cells through epigenetic reprogramming. A study of tumor and non-tumor epithelial cell lines revealed that their exposure to a stiff matrix induces migration driven by the activation of the YAP transcriptional program. The cells retained enhanced actomyosin expression and nuclear YAP translocation induced by a stiff matrix even after exposure to a soft secondary matrix, suggesting that matrix stiffness induces mechanical memory through epigenetic changes [348,349]. The high viscosity of the tumor interstitial fluid is associated with the leakage of the lymphatic vessels and increased degradation of ECM, leading to the accumulation of macromolecules [149]. This elevated interstitial viscosity induces TRPV4 activation and RhoA-mediated cell contractility. Breast tumor cells exposed to elevated viscosity exhibited increased migration and extravasation potential and high lung colonization in murine xenograft models. Tumor cells previously exposed to elevated viscosity retained high migratory potential after switching the medium viscosity to baseline level. This viscous cell memory was mediated by TRPV4 and Hippo signaling [149]. Thus, mechanical memory in the form of epigenetic adaptations induced by the primary tumor microenvironment can influence tumor cell survival during the entire metastatic journey and colonization of distant organs [149,349].
Tumor cells can spread from the primary site as single cells or clusters and can be detected in the bloodstream in the form of single or clustered CTCs [328]. A minimally invasive method of the CTC enumeration in peripheral blood samples called liquid biopsy can be used as a prognostic marker in patients with breast, prostate, and colorectal cancer [350]. It is based on immunomagnetic capture of cells positive for the epithelial cell adhesion molecule (EpCAM) in the blood samples. However, some CTCs are EpCAM negative as a result of their phenotypic plasticity, such as EMT [351]. This plasticity is induced by different microenvironmental stimuli, including mechanical stresses. The CTCs in the bloodstream are exposed to hydrodynamic shear stress (HSS), which can affect the metastasis-initiating properties of these cell populations. Indeed, HSS has been shown to trigger EMT phenotypes [159,352], and induce CSC properties through the inhibition of the extracellular signal-related kinase (ERK)/glycogen synthase kinase (GSK)3β signaling [353]. Shear stress induces a number of signaling pathways, including PI3K/Akt, MMPs, and YAP/TAZ [354–356], and promotes tumor-forming and migration potential [353,355,357]. Therefore, CTC exposure to the HSS in the bloodstream might induce their reprogramming toward more aggressive metastasis-inducing populations. In contrast to the highly invasive soft tumor cells, CTCs are mechanically robust [299], allowing them to withstand HSS. Some CTCs are protected from HSS-mediated damage by inducing RhoA signaling, increased formation of cortical F-actin, and activation of myosin II [299]. Despite the adaptive mechanisms, metastatic colonization is a rare event, and less than 0.01% of tumor cells entering the bloodstream give rise to macroscopic metastases [358]. Different microenvironmental factors, including HSS and cell compression upon squeezing through the wall of blood vessels and small-sized capillaries, might induce NER and cell death. On the other hand, mechanical deformation selects the tumor subpopulation with resilience to cell death induced by mechanical deformation. This cell population possesses highly activated DNA damage response, upregulated proliferation, and resistance against chemotherapeutic drugs [359]. Thus, the biomechanical characteristics of tumor cells are dynamically changing through epigenetic reprogramming or selection processes depending on the applied physical forces. This biomechanical plasticity provides tumor cells with high adaptability during tumor growth and metastatic dissemination in the dynamic and heterogeneous microenvironment [34,360] (Figure 6).
5.2.4 Electrical features of CSCs
Electric fields regulate the key biological functions of cells, including proliferation, stemness, and differentiation [361,362]. In addition to their distinct mechanical properties, CSCs have unique electrical characteristics. As discussed in Section 3, cancer cells possess unique bioelectric properties, making them distinct from their normal counterparts. The bioelectric properties of tumors and CSCs are defined by the above-mentioned Ca2+ ion channels TRP, Piezo as well as K+, Na+ and Cl+ channels, the intracellular and extracellular pH, and mitochondrial characteristics including pH, membrane potential, and ion concentrations [183,202,325,363,364]. Consequently, the seminal studies demonstrated the unique bioelectrical features of CSC cells.
A depolarization of the cell membrane was suggested as a marker of CSC populations [204], whereas induction of cell membrane depolarization by a suitable electrical environment was associated with CSC differentiation in glioblastoma (GBS) and was proposed as one of the therapeutic approaches to eliminate CSC populations [365]. The electrostatic potential difference (EPD) measured on cryosections of different tumor tissues correlates with tumor spread and CSC abundance, suggesting the prognostic potential of EPD measurement [366]. Furthermore, galvanotaxis study revealed that CSCs migrate to the regions with negative electrostatic potential [366].
A single-cell impedance measurement revealed that CSC and non-CSC populations from liver and oral squamous carcinoma cell lines can be differentiated within the impedance magnitude ranging between 2 and 20 kHz [283]. The stem cell phenotype of these cells was verified by the analysis of CSC biomarkers. Another study measured the bioelectrical impedance of non-cancerous breast epithelial cells and breast cancer cells with different metastatic potential in the range of 200 kHz-5 MHz. This study revealed a high correlation of the conductivity (σ) and permittivity (ε) to the tumor cell characteristics, including metastatic potential and migration capacities, and expression levels of different markers indicative of cell proliferation, microenvironment hypoxia and acidification, and glucose metabolism. Highly metastatic breast cancer cells have shown higher relative ε than non-metastatic tumor cells, and this difference is even more prominent compared to normal epithelial cells. These distinct electric properties can be attributed to the activation of the ion channel proteins such as Na+/H+ exchanger 1 (NHE1), which transports a proton to the extracellular space, high lactate secretion and, correspondingly, increased extracellular acidification [165]. Hence, this methodology can be used to monitor CTCs in the bloodstream as a prognostic marker of metastatic tumors. The importance of electrical signaling for tumor progression was demonstrated by finding that metastatic potential can be inhibited by non-contact induced electric fields (iEFs), suggesting a new treatment strategy to prevent tumor dissemination [367].
6. Application of Biophysical Properties in Clinical Trials
6.1 Imaging and targeting tumor stiffness
The complex interplay between the biological and physical properties of tumor cells and the microenvironment drives tumor progression. Physical forces play a critical role in cancer growth but also can be used for its treatment. Radiation therapy in the form of electromagnetic waves has been one of the mainstays of cancer treatment for almost 130 years since German scientist Wilhelm Conrad Röntgen described the discovery of X-rays in 1895 [368]. X-rays are electromagnetic waves inducing tissue ionization and leading to a chain of biochemical reactions, starting from molecular damage to the death of tumor cells. Radiation therapy affects cells directly through DNA damage and indirectly through radiolysis of water and the production of chemically reactive oxygen species (ROS). Today, radiotherapy is applied for about 50–60% of all cancer patients during their treatment [369,370]. X-rays are also broadly used for non-invasive imaging techniques such as computed tomography (CT) [371].
Although the translation of biophysics into clinics is relatively new, the increased stiffness of solid tumors has been known since ancient times. The mechanobiology-based method of tumor palpation is also a part of clinical practice for the initial tumor diagnosis. Imaging techniques such as MRI and ultrasonography (US), which enable the evaluation of tissue stiffness, are critical non-invasive diagnostic and prognostic tools to predict clinical outcomes [372–375]. To improve the specificity of MRI, advanced MR imaging techniques, including magnetic resonance elastography (MRE), have been clinically tested to detect tumor tissues in patients with liver [376], brain [374,377], breast [378] and prostate cancer [379]. The endoscopy-guided US, also called echoendoscopy, is routinely used in clinical practice for the visualization of the gastrointestinal (GI) tract and nearby tissues and is a standard imaging tool for tumor diagnostics and treatment application [380]. Both MRE and US were proved to be reliable techniques for differentiating benign and malignant lesions with high sensitivity and specificity [376,379–384]. The presence of dense, collagen-rich ECM around a tumor, called desmoplasia, is a typical pathological feature of different types of tumors, including breast cancer [385]. Breast tissue density measured with X-ray imaging (mammography) has been strongly associated with increased risk for breast cancer development [385,386]. Collagen deposition rises in response to cancer therapy, serving as a marker of treatment response and being associated with improved outcomes in patients with pancreatic ductal adenocarcinoma (PDAC) [387,388]. The positron emission tomography (PET) 68Ga probe based on the collagen-binding peptide (68Ga-CBP8) was successfully used in a clinical setting for specific targeting and non-invasive quantifying the dynamic changes of type I collagen in tumor ECM [389] (clinical trial number NCT04485286). This type of ECM-specific imaging is a promising tool for non-invasive monitoring treatment response in patients with PDAC and potentially other types of tumors. A high expression of integrins in many cancer types is employed to develop PET tracers based on the integrin-binding peptides. These PET tracers were clinically tested for breast, pancreatic, lung, colon, prostate, head and neck, and other tumors positive for αVβ3 integrin (e.g., clinical trials NCT05013086, NCT05976620, NCT02747290), αvβ6 integrin (e.g., clinical trials NCT05835570, NCT04285996, NCT03164486) [390], and αvβ8 integrin [391]. The binding sites for the above-mentioned integrins possess very similar surface-interface properties, potentially challenging the binding specificity of the PET tracers [392].
Given the critical role of stiffness in tumor initiation, progression, and therapy response, targeting ECM components and mechanosignaling is a promising anti-cancer strategy. One of the approaches to decrease of ECM stiffness is the inhibition of lysyl oxidase (LOX), the enzymes mediating crosslinking and stabilization of ECM. LOX are copper amine oxidases inducing covalent crosslinking collagen and elastin proteins by oxidizing their lysine residues [393]. LOX proteins are deregulated in several types of cancer, and their expression levels correlate with tumor metastases and therapy resistance [394,395]. The LOXL2-targeted humanized antibodies Simtuzumab (GS-6624) were tested in combination with chemotherapeutic drug gemcitabine for the treatment of metastatic pancreatic adenocarcinoma in phase II clinical trial (trial number NCT01472198). Although this combination treatment was tolerable, it failed to improve clinical outcomes in these patients [396]. The later-developed chemical drugs were reported to provide more efficient inhibition of LOX activity than antibodies [397,398]. The chemical inhibitors of LOXL2, PXS‐5338, PXS 5382A, and PAT-1251 entered early-phase clinical trials for analysis of their safety and tolerability in healthy subjects and in patients with myelofibrosis (clinical trial numbers NCT02852551, NCT04676529, NCT04183517, and NCT04676529). The administering of PXS‐5338 results in efficient inhibition of LOXL2 activity in blood plasma [397]. The recent preclinical study described selective and potent pan-lysyl oxidase inhibitor PXS-5505. Treatment with this drug decreased tumor growth and metastases, decreased chemotherapy-induced collagen crosslinking and stiffness of the tumor microenvironment in human patient-derived xenografts (PDX) and genetically engineered mouse models [399]. This inhibitor was reported to provide long-lasting, potent inhibition of LOX and to be safe in a phase 1 clinical trial in healthy human subjects [400]. Future clinical trials have to evaluate the clinical efficacy of this treatment in patients with malignant diseases.
Another promising approach is targeting mechanotransduction pathways, including inhibition of TRP ion channels and integrins. The expression of the TRPV6 calcium channel is increased in response to the mechanical tension and is associated with the development of different types of cancer [401,402]. The chemical inhibitor of TRV6, SOR-C13, was safe and tolerated, and demonstrated antitumor activity in a phase I clinical study in patients with advanced solid tumors (clinical trial number NCT00839631 and NCT01578564) [403]. Another receptor of the TRP family, the TRPV4 calcium channel, can be activated by diverse stimuli, including altered cell volume, shear stress, and temperature changes [404–406]. The TRPV4-induced signaling mechanisms are critical for tumor proliferation and metastases [407-409]. The TRPV4 inhibitor GSK2798745 was well-tolerated in phase 1 clinical study involving healthy volunteers and stable heart failure patients (clinical trial number NCT02119260)[410]. The cold- and voltage-induced calcium channel TRPM8 is deregulated in tumors. It plays divergent roles in the regulation of cancer cell proliferation and invasion depending on the tumor entity [411]. The results from a phase 1 study of D-3263 HCl, a TRPM8 agonist, in patients with advanced solid tumors have shown evidence of on-target activation and disease stabilization in patients with advanced prostate cancer (clinical trial number NCT00839631) [412].
Integrins play a critical role in mechanotransduction by linking ECM and cell cytoskeleton, mediating the signals from other mechanoreceptors, and converting mechanical stimuli into intracellular signaling regulating tumor cell survival, migration, and treatment response [413–415]. Since their discovery around four decades ago, different members of the integrin family have been investigated as biomarkers and pharmacological targets. To date, seven drugs targeting integrins have been approved for clinical use by the FDA for the treatment of cardiovascular diseases, multiple sclerosis, plaque psoriasis, inflammatory bowel disease, and dry eye disease [416]. Targeting integrins in cancer has not yet been translated efficiently into clinics. The phase 3 multicentre trial testing the selective αvβ3 and αvβ5 integrin inhibitor cilengitide in combination with temozolomide and radiotherapy in patients with glioblastoma did not reveal additional toxic effects from cilengitide but also did demonstrate clinical benefit from combination of cilengitide with temozolomide chemoradiotherapy (clinical trial number NCT00689221) [417]. Another study of the monoclonal antibody PF-04605412 directed against the α5β1 integrin in patients with advanced or metastatic solid tumors was terminated due to an unfavorable safety profile (clinical trial number NCT00915278). The phase II study tested the antitumor activity of the monoclonal humanized antibody MEDI-522a (etaracizumab) targeting αVβ3 integrin with or without dacarbazine chemotherapy in patients with metastatic melanoma (clinical trial number NCT00066196); however, the results have not been reported yet. Additional integrin-targeting therapies such as α5β1-specific inhibiting antibody volociximab [418], anti-αv antibody abituzumab [419] and intetumumab [420] and several other antibody-based integrin-targeted therapies are being tested in early-phase clinical trials. However, most of the completed studies in patients with cancer did not demonstrate significant clinical efficacy that could be partially explained by the complexity of integrin biology, the multifaceted role of integrins in cancer and non-tumor cells such as immune cells, and the lack of biomarkers to predict patients response to the anti-integrin treatment [416,421,422].
6.2 Electricity-mediated cancer therapy
As discussed previously in Section 4, tumor and normal cells possess different electric properties. Transdermal exposure of tumors to the 100-400 kHz AC electric fields is a non-invasive treatment called tumor treating fields (TTFields) applied by the transducer arrays placed on the patient‘s scalp close to the tumor. The anti-cancer effect of TTFields depends on different factors, including treatment durations, used frequency, direction of the fields, and their intensities (V/cm) [423]. TTFields therapy induced a broad spectrum of biological processes in tumor cells, including inhibition of cell proliferation and migration, increasing membrane permeability, inducing replication stress and autophagy, and activation of the immune response [424,425]. Since the treatment with TTFields targets highly proliferative cells, tumors can be more sensitive to this therapy than adjacent normal tissues [426–429]. The anti-proliferative effect of TTFields was attributed to its impact on the proteins with large dipole moments like tubulin dimers and microtubules. TTFields induces conformational rearrangement and depolymerization of tubulin microtubules critical in cell division [426,428,430]. TTFields also affect septins, the proteins with high dipole moment required for positioning the mitotic spindle and cytokinetic progression leading to the aberrant cytokinesis, cell cycle block, and cell death [426,431]. The clinical benefits of anti-cancer therapy with TTFileds were mostly extensively analyzed in patients with glioblastoma (GBM) [432]. The electric field therapy was well tolerated in patients with GBM. The TTFields therapy in combination with temozolomide chemotherapy was more clinically efficient than therapy with temozolomide alone, which was evidenced by significant improvement in overall survival and progression-free survival (clinical trial number NCT00916409) [433,434]. The efficacy of the TTFields in clinical trials led to its FDA approval for patients with GBM in 2015. Several other clinical trials for TTFields therapy in combination with chemotherapy have shown its clinical benefit without an increase in toxicity for patients with mesothelioma (clinical trial number NCT02397928) [435], advanced non-small cell lung cancer (NSCLC) (clinical trial number NCT00749346) [436], advanced pancreatic cancer (clinical trial number NCT01971281) [437] and some other tumor entities [424]. These clinical data suggest that TTFields therapy has great potential to improve the outcomes of patients diagnosed with different types of malignancies. A broad variety of underlying molecular mechanisms enable its combination with different types of anti-cancer treatment [424,425,432].
The second clinically approved electricity-assisted therapy is irreversible electroporation (IRE). An external IRE increases cell membrane permeability. The irreversible membrane structural defects induced by IRE make cells more permeable to chemotherapeutic drugs, induce tumor cell death, and activate anti-tumor immune reponses [438,439]. IRE is performed under the US or computer tomography guidance using the NanoKnife IRE generator system [439,440]. Promising therapeutic effects and safety profile resulted in the clinical approval of The NanoKnife IRE system for ablation of soft tissue tumors. The PRESERVE study for the use of IRE treatment in patients with prostate cancer (clinical trial number NCT04972097) confirmed favorable treatment safety. The final results of the study are required to evaluate the treatment effectiveness [441]. NanoKnife IRE treatment is currently analyzed alone or in combination with conventional therapies in several additional clinical trials for different types of malignancies, e.g., pancreatic cancer (clinical trial numbers, e.g., NCT02791503, NCT03105921, NCT03180437), colorectal liver metastases (clinical trial number NCT02082782), breast cancer (clinical trial number NCT02340858), stomach tumors (clinical trial number NCT02430636), bladder cancer (clinical trial number NCT02430623) and others. The published results of these studies confirm the safety and the evidence of anti-tumor efficacy of the NanoKnife IRE therapy [442–445].
6.3 Thermotherapy
Thermoregulation is an evolutionarily developed adaptive mechanism enabling mammals to survive environmental alterations. Consequently, the biochemical reactions in the cells of mammalian organisms are highly temperature-sensitive [446,447]. A tissue heating of just a few degrees induces protein denaturation and aggregation, inhibits protein synthesis and DNA repair, and leads to cell cycle arrest and cell death [446,448]. Cancer cells are more sensitive to temperature deviations than normal cells. Depending on the applied temperature, local heating affects tumor access to a blood supply, and, therefore, availability of oxygen and nutrients; downregulates DNA repair and replication enzymes and induces DNA damage; impacts the tumor microenvironment, and activates anti-tumor immune response [449]. Thermotherapy, or thermal ablation, is an exposure of tumor tissue to temperature conditions different from its physiological diapason between 36 and 37.5 ℃. Thermotherapy is classified as hyperthermia (applying increased temperature of >40 ℃) or hypothermia (applying decreased temperatures of <–40 ℃) [450,451]. The anti-cancer effect of hypothermia has been shown for preclinical models of tumors with mutated p53 tumor suppressor [452], but in contrast to the hyperthermia, it was not developed into conclusive clinical trials for cancer treatment. Tumor heating can be achieved by different techniques, including electromagnetic fields with different frequencies, wavelengths, and tissue penetration (including radiofrequency waves of 0.3–30 MHz or microwave systems powered by 433 MHz, 915 MHz or 2,450 MHz generators) [453,454]; US heating using acoustic waves with a frequency on 0.5-10 mHz, infrared heating with frequency >300 GHz, laser light or perfusional hyperthermia or local heating techniques through physical contact with heating courses [453,455]. Thermotherapy is a non-invasive and safe therapy with clinically proven anti-cancer activity when used in combination with other treatments. Due to its effect on the blood flow, intratumor hypoxia, and tumor microenvironment hyperthermia is a potent sensitizer of tumor tissues to other anti-cancer treatments such as radiotherapy, chemotherapy, and immune therapy [450,456,457]. Thermal therapy techniques have been clinically approved for the management of several malignant diseases. A nanoparticle-mediated hyperthermia treatment using superparamagnetic iron oxide nanoparticles (SPIONs) subjected to alternating magnetic fields (AMF), and has been approved by the European Medicine Agency (EMA) for glioblastoma treatment [458]. Several clinical trials have evaluated the efficacy of hyperthermia alone or in combination with chemotherapy and radiotherapy [459–461]. Most of the studies reported evidence of the clinical benefit from hyperthermia treatment and similar rates of toxicity for the control arm (conventional therapy) and experimental arm (thermotherapy and conventional therapy), and many other trials are currently in progress for different tumor types, including head and neck (clinical trials number NCT00848042), prostate cancer (clinical trials number NCT02680535), and locally advanced cancers (clinical trials number NCT05099809) [459,462–464]. The clinical studies suggest that thermotherapy treatment holds great promise for improving cancer treatment in combination with conventional therapies. Nevertheless, although hyperthermia has been known from ancient times, the mechanisms of its action are still not wholly understood, and much work is required to understand the intracellular and tissue processes mediating the effects of hyperthermia and its possible synergism with other treatment modalities and define the optimal treatment protocols [453,457].
7. Predicting Cancer Outcomes with Biophysical Tumor Properties and Artificial Intelligence
Employing machine learning (ML) algorithms to predict clinical outcomes has a longstanding history; some examples include [465,466]. However, conventional ML algorithms like support vector machines require an additional step of manual feature engineering, i.e., qualitative or quantitative description of the diagnostic data. This limits both the scalability and applicability of the conventional ML algorithms to unstructured molecular or imaging data. In contrast, the current generation of AI algorithms is leveraging an approach called representation learning, allowing them to learn features directly from unstructured data algorithmically. One of the starkest examples of representation learning algorithms instrumental in predicting cancer outcomes is the application of convolutional neural networks to biomedical imaging of cancer (reviewed in [467]), such as in digital pathology [468] and MRI [469].
Crucially, management of cancers like lung, brain, breast, and prostate, where traditional radiographic assessment may be limited, has been shown to improve by applying representation learning algorithms [469]. Specifically, since tasks like the demarcation of tumor volume and the extraction of characteristic cancer phenotypes can be addressed more consistently, integrating AI-based workflows contributes to the improvement of risk prediction and, ultimately, outcomes. Furthermore, in the case of digital pathology, representation-learning algorithms allow for rapid analysis of large whole-slide images by detecting tumor regions (image segmentation) and categorizing the tumor microenvironments by identifying their respective cellular composition (instance segmentation) [468,470,471]. Notably, these approaches can lead to new ways of characterizing tumor microenvironments in pathology images through determining factors like tumor purity. In a recent study, Gong et al. have shown that low tumor purity is associated with unfavorable outcomes and immune evasion phenotypes in gastric cancer, highlighting the importance of microenvironment characterization [472].
Beyond imaging, recent advances in representation learning include unstructured multi-omics data, which allows for the leveraging of genetic and molecular profiles, as well as clinical data as predictive hallmarks [473]. For example, AI algorithms used in natural language processing have been successfully employed to facilitate the annotation of multi-cancer genomic datasets [474]. Finally, representation learning techniques allow for joint training of multi-modal models on diverse data with cancer prognosis as the objective [475].
Furthermore, representation learning has been applied for the analysis of complex data in the field of translational biophysics, for example, for the identification of tumor cells, classification of their phenotype, and therapy response based on their electrical properties measured with EIC [164,476–478], for correlation of tumor stiffness and cancer treatment efficacy [479,480] or prediction of patients’ risk of metastases based mechanobiological assays [481].
8. Challenges and Perspectives
The physical and biological properties of tumor cells are tightly interconnected and mutually dependent. Increasing ECM stiffness and hypoxia are the fundamental hallmarks of tumor progression and poor therapeutic response. Together with other associated tumor features such as metabolic programming and lactic acidosis, genomic instability, and upregulation of the oncogenic pathways, including ion channels, they not only fuel tumor aggressiveness and therapy resistance but also alternate the physical parameters of tumor cells and microenvironment, including electrical, mechanical, and thermal properties. Across different solid tumor entities, there is wide accordance that tumors show impedimetric parameters different from normal tissues and characteristic mechanical changes at the intracellular, cellular, ECM, and interstitial fluid levels that bear great potential for new biomarker developments. Recent preclinical developments demonstrated that multiple physical and mechanical properties might serve as promising diagnostic, prognostic, and predictive biomarkers. Still, many questions remain about how to facilitate their clinical translation. A current limitation is the comparatively low number of studies that exploit reasonable numbers of patient-derived materials for single-cell and intracellular experiments. In order to develop clinically applicable routines, a higher level of standardization of measurement protocols, larger patient cohorts, longitudinal sample analysis, and higher throughput of measurements are needed. In addition to probing cancer cells themselves, more future studies may also be expanded on other cell types of the tumor microenvironment, e.g., on stromal cells [482,483] and immune cells. Furthermore, measurements of cells from solid tumors typically require digestion and dispersion of tissues to get access to single cells [47]. On the other hand, testing of patient-derived cancer cells from pleural fluids or blood from liquid biopsy samples does not require enzymatic digestion and can better reflect the native mechanical properties of analyzed cell populations.
The development of improved preclinical models that can recapitulate tumor physical parameters and biological clues is one of the main challenges for translational research. The earlier studies focused on the tumor biophysical properties by employing 2D cell culture. However, these 2D models did not recapitulate the complexity of the tumor interaction with the microenvironment. This concern is partially addressed using 3D in vitro culture, where tumor cells alone or together with stroma cells are embedded in the synthetic and natural biomatrix such as hydrogels or animal-derived matrigel. Although these models better resemble tumor architecture, they fail to fully recapitulate the physical forces exerted on cells in tissues. The recently developed technology, such as organ-on-a-chip, combines tumor cell culture along with ECM components with fluid flow. These models are used to recapitulate tumor mechanobiology better and investigate the role of mechanical forces in the regulation of tumor cell survival, invasion, and therapy resistance [34,484].
Although the thermal properties of cells exhibit significant changes between healthy and cancerous states, highlighting their potential as prognostic biomarkers, the thermal methods have not yet garnered the same level of attention as electrical or optical techniques in biomedical research and diagnostics. This disparity can be attributed to several challenges, particularly in the experimental setup. Achieving high sensitivity in thermal measurements requires effective thermodynamic shielding from environmental influences, a challenge seen in methods such as calorimetry and HTM method. Despite these limitations, the simplicity of the read-out principles and the potential for indirect and label-free sensing in thermal techniques offer promising avenues for future exploration. With advancements in experimental design and improved sensitivity, thermal methods could emerge as a valuable tool in cellular analysis and cancer prognosis. Further research and innovation are needed to overcome current barriers and fully realize their potential.
Functional and phenotypical diversity of cancer cells within individual tumors is one of the major challenges for efficient tumor treatment. Tumors are highly heterogeneous tissues consisting of different types of cells, with different potentials to initiate and maintain tumor growth, withstand therapies, and metastasize. The eradication of all tumor-maintaining CSC cells is critically required for tumor treatment. Despite stemness being more transient than stable cell populations, CSCs are the critical determinant of tumor growth during cancer progression and its regrowth after treatment [279,485]. Thus, eradication of all CSCs is essential for permanensst tumor control [486]. However, the biological markers of CSC often lack specificity and are expressed on other types of cells. Due to the lack of precise tools to identify CSCs, the targeting of these populations still remains elusive. The maintenance of CSC in their niches depends on the mechanoenvironment. The intra- and intertumoral mechanical heterogeneity of tumor tissues impacts the distribution and properties of CSCs. The maintenance of CSCs in their niches depends on the microenvironment. The intra- and intertumoral mechanical heterogeneity of tumor tissues impacts the distribution and properties of CSCs. Thus, targeting the mechanical microenvironment can be a promising target for eradicating CSC populations [487]. Furthermore, spatial mapping and longitudinal analysis of the mechanical and electrical properties of CSCs can bring an opportunity to develop novel label-free prognostic markers. Analysis of the physical characteristics of CSCs and their correlation with biological, pathological, and clinical parameters can better explain tumor heterogeneity and help in tailoring cancer treatment to individual patients.
Although the biological mechanisms making tumor cells more sensitive to the electricity- and temperature-driven treatment are not fully understood, several clinical studies confirmed the efficacy and safety of hyperthermia and electricity-assisted anti-cancer therapy. Still, much work is required to decipher the comprehensive biological mechanism activated by these treatments in tumors and surrounding tissues and to develop predictive markers for the patient’s response. To establish a meaningful correlation between complex imaging, molecular and electrical tumor parameters, and therapeutic responses or clinical outcomes, ML models are currently used. The AI and improved computing power are making possible to connect biomarkers with biophysical properties and clinicopathological parameters to develop novel label-free diagnostic and prognostic tools.
Supplementary Materials
The following supplementary materials are available on the website of this paper: CHP2401020011SupplementaryMaterials.zip
Table S1. Most important parameters that can be extracted from impedance and how to derive them.
Declarations
Competing Interests
Anna Dubrovska is a member of the Editorial Board of the journal Cancer Heterogeneity and Plasticity. The author was not involved in the journal’s review of or decisions related to this manuscript. The author has declared that no other competing interests exist.
Acknowledgement
J.-F. B. thanks G. Arkowitz, B. Ladoux, R.-M. Mège and M. Reffay for stimulating discussions. This research was supported by the Agence Nationale de la Recherche (ANR) under the contract ANR-24-CE42-6142-03 (VISCOMAG2) (to J.-F. B.) and by the HZDR-Programm für Vernetzungsdoktorand*innen (to L.B. and A. D.).


 ,
Daniel Nieder
2
,
Daniel Nieder
2