1. Introduction
Bladder cancer (BC) is the fourth most common cancer in males in the United States, with an estimated 83190 new cases and an estimated 16840 deaths in 2024 [1]. Almost 90% of bladder cancers are urothelial carcinoma (UC). Approximately 25–30% of muscle-invasive bladder cancers (MIBC) exhibit divergent differentiation, such as squamous, glandular, adenocarcinoma, micropapillary, plasmacytoid, trophoblastic, and neuroendocrine (NE), etc. [2]. According to the WHO classification of Urinary and Male Genital tumors, neuroendocrine bladder cancer (NEBC) represents ~1% of urinary BC, including small cell neuroendocrine carcinoma (also known as small cell bladder cancer, SCBC), large cell neuroendocrine carcinoma (LCNEC), well-differentiated neuroendocrine tumor, and paraganglioma [3,4]. SCBC represents the most common NEBC [3,5]. The first reported case of SCBC was described by Cramer et al. in 1981 [6]. SCBC is a rare and highly aggressive cancer with a poor prognosis and a lack of standardized treatment options. The therapeutic management of SCBC is empirically guided by clinical strategies and insights derived from small cell lung cancer (SCLC) [7–9]. The rarity of SCBC limits genomic, biological, and clinicopathological research, leaving patients with limited treatment options beyond the standard chemotherapy regimen of Etoposide and Cisplatin. Despite significant advances in the basic understanding over the past decades, SCBC remains understudied, with challenges in uncovering its pathogenesis, tumor heterogeneity, plasticity, cell of origin, and potential therapeutic targets.
Numerous studies have reviewed the epidemiology, clinical presentation, diagnosis, treatment, and prognosis of SCBC. Identified risk factors for SCBC include smoking, male sex, and advanced age [10]. SCBC is observed five times more frequently in men than in women, with cigarette smoking history in 50–70% of patients [11–13]. The average age of onset for SCBC is 72 years [14]. The most common symptom is painless gross hematuria (67–100% in SCBC) with or without dysuria [15]. Other symptoms include urinary obstruction, abdominal pain, pelvic pain, recurrent urinary tract infection, or weight loss [11,16–18]. The diagnosis of SCBC is based on histopathological examination of tumor tissue obtained by cystoscopy and transurethral resection of the bladder tumor (TURBT) or radical cystectomy [19]. The current clinical management of SCBC includes chemotherapy as the first approach, along with radical resection, radiotherapy, or multimodality treatment strategies [10,20]. Second-line immunotherapy demonstrates limited efficacy in the treatment of SCBC [21]. The overall prognosis for SCBC remains extremely poor. The median survival of SCBC patients ranges from 12 to 24 months with treatment, but only 4 to 5 months without treatment. The five-year survival rate varies between 8% and 40% [10,17,18,22]. Like SCBC, LCNEC is frequently presented with locally advanced and high-stage disease [23,24]. LCNEC has been treated using a similar approach to SCBC, involving neoadjuvant chemotherapy (NAC) followed by cystectomy [5]. In a cohort of 43 LCNEC patients and 192 SCBC patients, LCNEC patients demonstrated longer overall survival (OS) compared to those with SCBC only [25]. However, in another larger cohort of LCNEC (n = 80), LCNEC showed the worst disease-free survival compared to SCBC or mixed NEBC tumors in patients who underwent cystectomy [26].
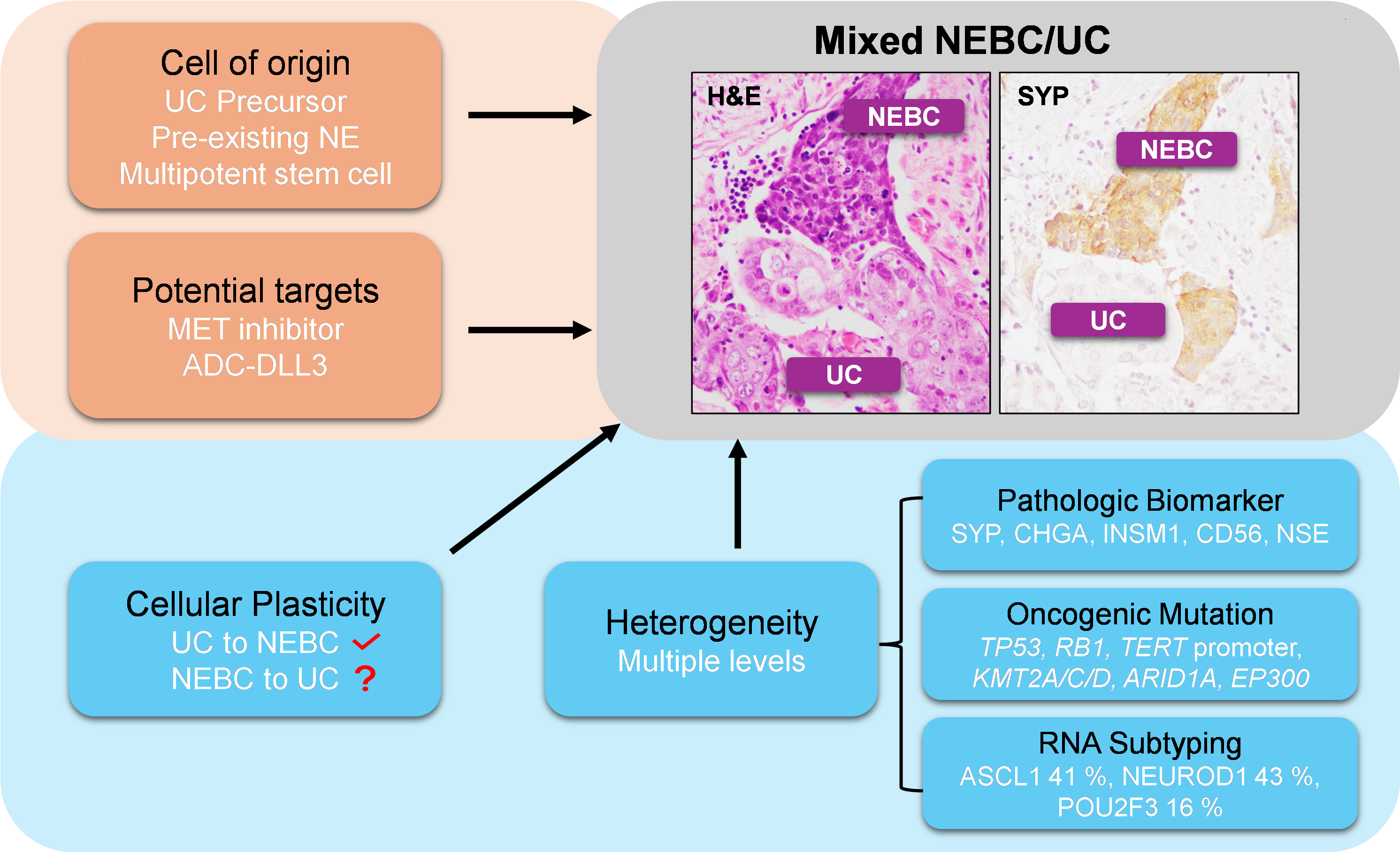
In this review, we summarize the molecular characteristics of NEBC and discuss its underlying tumor heterogeneity, cellular plasticity, cell of origin, and potential targets of NEBC or NE-like tumors (Figure 1). This review aims to enhance understanding of NEBC tumorigenesis and classification, guide the discovery of new therapeutic targets, and improve the effectiveness of treatment approaches.
2. Histopathology and Immunohistochemistry of NEBC
The histological features of SCBC are almost identical to SCLC, sharing the same diagnostic criteria established by the WHO classification system [20]. Morphologically, SCBC exhibits small cells with a high nuclear-to-cytoplasmic ratio, nuclear molding, abundant mitotic figures, and necrosis. Immunohistochemical (IHC) staining with NE biomarkers, including chromogranin A (CHGA), SYP, CD56, neuron-specific enolase (NSE), and insulinoma-associated protein 1 (INSM1), assists in the diagnosis of NEBC [13,27,28]. Due to the 38–70% coexistence of SCBC with UC and its differentiated components, epithelial biomarkers such as cytokeratins, p63, p40, EMA, and CAM 5.2 are also utilized in its diagnosis [12,18,20,29]. Compared to SCBC, LCNEC comprises large, high-grade polygonal tumor cells that exhibit variable amounts of cytoplasm and prominent nucleoli [2]. LCNEC is also characterized by typical NE markers including CD56, synaptophysin, chromogranin, and INSM1 [30]. Figure 2 illustrates the histological features of three SCBC patients alongside their corresponding patient-derived xenografts (PDXs).
3. Genomic Alterations in NEBC
Next-generation sequencing has provided valuable insights into the mutational profiling of NEBC, offering genetic evidence to investigate its pathogenesis. SCBCs exhibit significant pathologic and genomic similarities to SCLC. Table 1 summarizes the common genetic alterations in NEBC. Like SCLC, SCBC frequently harbors alterations in TP53 and RB1, with a high prevalence of co-alterations in both genes [7,31]. However, the TERT promoter was identified as a bladder-specific mutation with high frequency, which is absent from small cell carcinoma of other origins including lung [7,32,33]. The most frequent mutations in LCNEC include TP53 (92%), RB1 (65%), and TERT promoter (76%). Additionally, LCNEC harbors potentially targetable alterations in ERBB2, ERBB3, and PIK3CA at a frequency of 11% [26]. In another cohort of 18 LCNECs, mutations were identified in the TERT promoter (89%), TP53 (100%), and RB1 (100%), with 56% of tumors exhibiting co-alterations in TP53 and RB1 [25].
In a cohort of 61 SCBC patients from Memorial Sloan Kettering Cancer Center (MSKCC), the most frequently altered genes were TP53 and RB1, found in 90% of patients, with co-alterations occurring in 80% [7]. A comparison study was conducted to identify similarities and differences in histology, cell lineage, and organ specificity between SCBC and SCLC tumors. The whole-exome and/or MSK-IMPACT sequencing identified a high prevalence of APOBEC-mediated mutational processes in SCBC, revealing a distinct pathogenic mechanism driving smoking-associated SCBC compared to SCLC. SCBC exhibited a significantly higher rate of biallelic mutations in TP53 and RB1 than UC. The co-existence of identical TP53 and RB1 mutations in UC suggests that these alterations alone are insufficient for the development of SCBC. SCBC-specific TERT promoter mutations were identified in 95% of patients but were absent in SCLC [33]. Moreover, mutations in epigenetic modifiers such as KMD6A, ARID1A, CREBBP, EP300, and KMT2A/C/D were identified in 74% of SCBC cases, resembling those found in UC but rarely observed in SCLC. These findings suggest that SCBC likely originates from a UC precursor. Evolutionary heterogeneity analysis of SCBC indicated that TP53 and RB1 are not the drivers of SCBC transformation from UC, but likely arose early in the molecular time, shortly after founding driver mutations. A subsequent genomic study from MSKCC analyzed 47 pre-treatment SCBC tumors in the neoadjuvant setting and correlated genomic alterations with treatment responses. TP53 (87%), TERT promoter (83%), RB1 (79%), KMT2D (28%), ERCC2 (26%), CREBBP (23%), ARID1A (19%), EP300 (15%) were altered frequently in SCBC [34]. Notably, ERCC2 mutations were significantly enriched in patients who achieved complete responses following NAC, consistent with the findings observed in UC [35,36].
Another recent genomic study from MD Anderson Cancer Center analyzed 34 SCBCs and 84 conventional UCs along with a reference of 408 MIBCs in the TCGA (The Cancer Genome Atlas) database [37]. The mutational profiles revealed that TP53 (93%) and RB1 (47%) were mutated in SCBC at a higher frequency than in UC from the TCGA cohort. SCBC and UC tumors shared several significantly mutated genes, particularly in paired cases containing both SCBC and UC components. Identical mutations in p53 (p.H179Y, p.C176F) and RB1 (p.R798fs) observed in paired tumors suggested a clonal evolutionary process with a shared origin. Moreover, no SCBC tumors exhibited the typical mutations enriched in luminal conventional UC, suggesting that SCBC likely originates from a basal lineage.
In a larger study, 3.92% (132) of SCBC cases were identified among 3,368 UC samples. The three most frequent mutations were TP53 (92%), RB1 (75%), combined TP53/RB1 (72%), and TERT promoter mutations (68%) [32]. Transcriptome RNA sequencing analysis of 24 pure SCBCs and 51 MIBC UCs revealed that SCBC expressed lower levels of canonical T-cell biomarkers, showed similar signature enrichment to SCLC, but displayed distinct gene expression profiles compared to UC. Similarly, a recent study of 44 SCBC cases (36 pure SCBCs and 8 mixed cases with small-cell components) identified frequent alterations, including TP53 (75%), RB1 (38%), and TERT mutations (92%), along with other mutations such as ARID1A (38%), MYCL (33%), and PIK3CA (21%) [38]. Another NEBC study included 103 SCBC and 19 LCNBC [39]. In 33 sequenced tumors, mutations in TP53, RB1, and TERT were detected in 30 (91%), 26 (79%), and 27 (82%) tumors, respectively. However, immunohistochemistry (IHC) results on the overall cohort indicated 93% (101/108) abnormal p53 staining and 92% (103/112) loss of Rb expression. The staining patterns of p53 and Rb did not show a significant difference in a paired analysis comparing NE and non-NE in the same tumor.
4. NE-like or Neuronal as a Subtype of MIBC
The ability to accurately differentiate BC subtypes can offer valuable prognostic insights and potentially predict therapeutic responses for precise patient stratification and personalized treatment approaches. Several subtype classification systems for MIBC have been reported over the past decade. The first molecular subtyping of MIBC was performed by a research group at Lund University [46]. Another research group at the University of North Carolina (UNC) used consensus clustering to divide MIBC into luminal and basal two subtypes [47]. Then, the Lund University group developed a newer classification system including five subtypes: urobasal A, genomically unstable, urobasal B, squamous cell carcinoma-like (SCCL), and an infiltrated class [48]. The same Lund group analyzed a cohort of 307 advanced bladder cancers by global gene expression, resulting in six consensus clusters [40]. Compared to the other known MIBC molecular subtypes clustering including UNC classification [47], MDA classification [49], and TCGA consortium [50], the six clusters were classified as urothelial-like, genomically unstable, epithelial-infiltrated, SCCL/mesenchymal-infiltrated, SCCL/UroB, and small-cell/neuroendocrine-like. Combined with IHC profiling, the cohort was divided into 5 tumor-cell phenotypes with molecular pathological definitions: urothelial-like, genomically unstable, basal/SCCL, mesenchymal-like, and small-cell/neuroendocrine-like [40]. This combination of molecular pathology (tumor-cell phenotype) and global mRNA profiling has been proposed for accurate subtyping of MIBC. This approach, known as the Lund taxonomy, was subsequently validated in a TCGA cohort [51].
A comprehensive analysis of the TCGA cohort of 412 MIBC cases identified five expression subtypes including luminal-papillary (35%), luminal-infiltrated (19%), luminal (6%), basal-squamous (35%), and neuronal (5%) [41]. Neuronal subtype includes 3 tumors with NE histology and 17 tumors without NEBC histologic features. All these tumors showed relatively high expression of neuronal differentiation genes and typical NE markers. However, the majority of these “neuronal” were not consistent with NEBC histologic features but were like conventional UC phenotypically, even though they have poor outcomes similar to NEBC.
Batista et al. analyzed transcriptome expression profiles of MIBC from seven institutions and developed a strict single-sample NE-like classifier by machine learning to identify NE-like tumors regardless of their pathologic presentation [43]. This classifier included an 84-gene panel and proved to be more stringent than previous subtyping models for NEBC. To validate the accuracy of the NE-like classifier, seven MIBCs with SCBC histology were 100% identified as NE-like. The study also utilized the CellMiner analysis tool to generate drug response scores, providing a numerical metric to predict chemotherapy responses for each NE-like tumor. NE-like tumors are predicted to be more sensitive to chemotherapy drugs, such as cisplatin, gemcitabine, and etoposide (agents commonly used in MIBC treatment), compared to basal and luminal tumors. A subsequent study validated this NE-like classifier and its ability to predict poor prognosis in a retrospective multicenter cohort of 234 patients treated with cisplatin-based NAC [52]. Ten patient samples were classified as the NE-like subtype with by strong expression of NE markers and the absence of basal or luminal markers. These patients exhibited significantly higher cancer-specific mortality rates compared to those with non-NE-like subtypes.
A comprehensive profiling study integrated six published MIBC classification systems from 1750 samples to develop a unified consensus subtyping framework. This system includes six subtypes: luminal papillary (24%), luminal non-specified (8%), luminal unstable (15%), stroma-rich (15%), basal/squamous (35%), and NE-like (3%) [44]. With the help of a single-patient classifier [43], this system enables the assignment of a consensus class to each patient, potentially facilitating more precise treatment strategies in the future. In this system, the NE-like class exhibited overexpression of neuroendocrine differentiation genes, lacked immune infiltration markers, was associated with the poorest prognosis, and displayed profiles suggestive of a potential response to radiotherapy. Concurrent TP53 (94%) and RB1 (94% present either mutation or deletion) inactivation were observed in the NE-like class. Notably, this classifier is limited by the presence of heterogeneous tumors, which may contain multiple subtypes. Further studies are needed to evaluate the impact of intratumor heterogeneity on its accuracy and applicability.
5. Molecular Subtyping of NEBC
Koshkin et al. analyzed distinct gene expression patterns in SCBC and corresponding normal bladder tissues [45]. Gene expression profiling of 40 tumors plus 6 “normal-appearing” tissues from histologically confirmed 63 SCBC cohort revealed 4 distinct subsets (cluster 1–4) by unsupervised hierarchical clustering. These clusters are associated with discrete clinical phenotypes that correlated with 5-year OS. Cluster 2 (“normal-like”) exhibited the best OS, while Cluster 3 (“metastasis-like”) was associated with a higher likelihood of metastasis and shorter OS (6 months). In multivariate analysis, DLL3 and CD56/NCAM1 expression in tumors were identified as negative prognostic biomarkers for SCBC. The efficacy of DLL3-targeting antibody-drug conjugates (ADCs) was validated in an SCBC PDX model, highlighting DLL3 as a potential therapeutic target in SCBC.
Using the cluster analysis for lineage-specific transcription factors (TFs) identified in SCLC [53,54], 44 SCBC cases were classified into three subtypes (ASCL1, NEUROD1, and POU2F3) and validated their respective downstream targets [38]. Specifically, mixed cases were enriched in the POU2F3 subtype. Heterogeneity in the immune marker expression in these subtypes may suggest their different response to immunotherapy. The NE markers expression-low POU2F3 subtype expressed the highest immune signature scores and the NE markers expression-high ASCL1 subtype significantly expressed CEACAM5, suggesting a potential response to CEACAM5-specific antibody-drug conjugate (ADC) - labetuzumab govitecan, currently under clinical investigation.
Recently, IHC for ASCL1, NEUROD1, and POU2F3 has been integrated into NEBC subtyping. Based on the analysis of the IHC co-expression pattern of ASCL1, NEUROD1, and POU2F3 in 116 SCBC/LCNBC tumors, 5 subtypes were identified as ASCL1+ (34%), NEUROD1+ (23%), ASCL1+/NEUROD1+ (16%), POU2F3+ (21%), ASCL1–/NEUROD1– (6%) [39]. These molecular subtypes have no specific microscopic H&E features to be distinguished reliably by histomorphology. Most POU2F3+ subtype tumors lacked traditional NE markers expressions (SYP, CHGA, INSM1) and showed reduced CD56 expression, in contrast to the ASCL1 and NEUROD1 subtypes. Furthermore, DLL3, a therapeutic target for antibody-drug conjugates or bispecific T-cell engagers, is highly expressed in 82% of SCBC cases. POU2F3+ and PLCG2+ (a top upregulated gene in SCLC associated with pro-metastatic and stem cell-like features [55]) are significantly associated with worse patient outcomes in the radical cystectomy cohort.
6. Tumor Heterogeneity in NEBC
Tumor heterogeneity plays an important role in tumor progression, resistance to treatment, and metastasis [56–58]. The complexity of heterogeneity includes clonal expansion of individual mutations, genomic alterations, and expression differences within different patient tumors (interpatient heterogeneity), different regions in one tumor (intratumoral heterogeneity), primary tumor and its metastatic tumor (intertumoral heterogeneity), and over time changing tumor (temporal heterogeneity) [59,60]. MIBC is a highly heterogeneous disease [61]. However, within MIBC, tumor heterogeneity in NEBC has not been systematically investigated.
At the level of histological morphology, within SCBC [62] or LCNEC [24], clinicopathologic differences were observed between tumors, including histopathology and immunohistochemistry, as well as incidence differences of mixed UC patterns and tumor stages differences. A heterogeneous immunophenotypical profile of urinary NEBC has also been observed [63]. Based on the differential IHC co-expression patterns of ASCL1, NEUROD1, and POU2F3 in 116 NEBC tumors, five distinct NEBC subtypes were identified: ASCL1+, NEUROD1+, ASCL1+/NEUROD1+, POU2F3+, ASCL1–/NEUROD1– [39]. Figure 2 also demonstrates mixed UC/SCBC histologic characteristics in 1 out of 3 SCBC patients.
At the mutational level, although the three most frequent mutations (TP53, RB1, and TERT promoter) are common, individual tumors from patients exhibited distinct mutation patterns. RB1 mutations are a common hallmark of SCBC; however, some SCBC patients lose RB protein expression despite the absence of detectable RB1 mutations [7]. Mutations in epigenetic modifiers, such as KMT2D, CREBBP, ARID1A, and EP300 were frequently altered [7,34]. However, these chromatin-remodeling genes were not frequently altered in another SCBC cohort [37]. ERCC2 mutations in some SCBC tumors are associated with improved responses to NAC [34].
At a transcriptomic level, molecular subtyping of SCBC has been studied for patient stratification. The distinct gene expression patterns within SCBC divided tumors into 4 distinct clusters [45], and the "metastasis-like" cluster was associated with shorter OS. A recent study grouped SCBC cases into three subtypes (ASCL1, NEUROD1, and POU2F3) using cluster analysis for lineage-specific TFs [38]. The POU2F3 subtype with low expression of NE markers exhibited the highest immune signature scores. Overall, the heterogeneity in NEBC remains understudied. A deeper understanding of NEBC heterogeneity could aid in developing more precise therapies, ultimately improving patient outcomes.
7. Cellular Plasticity of NEBC
Lineage plasticity refers to the ability of cells to reprogram and change to a different phenotypic identity [58,64]. These changes often involve transcriptional or epigenetic fluctuations, and occasionally genetic alterations, which lead to cell state plasticity to promote tumor initiation, progression, metastasis, immune evasion, and chemoresistance [65,66]. Few studies have investigated lineage plasticity in NEBC, but existing research suggest possible UC-to-NEBC plasticity. No conclusive evidence of NEBC-to-UC plasticity has been reported.
A comprehensive genomic profiling analysis of genitourinary NE (11 SCBC and 1 prostate small cell carcinoma resected from the bladder) was conducted [42]. Consistently, frequent alterations of TP53, RB1, PIK3CA, ERCC2, ARID1A, and EP300 were detected in SCBC, which were detected in UC as well [67,68]. Integrated analysis revealed three key altered function modules, including the p53/Rb pathway, receptor tyrosine kinase signaling, and epigenetic regulators. Together with the related works [7,69,70], NE and UC were proved to share generally identical genotypic and etiological characteristics, which supported that they share a common cell origin. Furthermore, preliminary results from CRISPR experiments targeting TP53 and RB1 in UC cell lines TCCSUP and J82 revealed an increase in multiple canonical NE markers, suggesting that the loss of TP53 and RB1 may drive a UC-to-NEBC lineage plasticity. This NE lineage plasticity has been also observed in EGFR mutant lung adenocarcinoma [71] and neuroendocrine prostate cancer (NEPC) [72,73].
Another study investigating microRNA-145 in human UC cell lines T24 and KU7 provided evidence supporting cell plasticity within UC tumors [45]. The overexpression of microRNA-145 increased stem cell markers and upregulated markers of squamous, glandular, and NE cells, suggesting that microRNA-145 could reprogram UC cells into multipotent stem-like cells, which subsequently differentiate into other lineages, including squamous, glandular, or NE. However, these studies are limited to in vitro molecular analyses and lack in vivo validation through NE marker staining and/or histological confirmation of NEBC morphology.
Wang et al. reported that EPCAM+/CD49f+ UC cells from the primary human renal pelvis or bladder were transformed into SCBC features by transducing a dominant-negative form of TP53, myr-AKT1, RB1 shRNA, C-MYC, and BCL2, collectively termed PARCB [74]. This artificial SCBC tumor displays diverse histological features, including SCBC, UC, and squamous differentiation. Single clones from bladder SCBC tumors exhibited both SCBC and UC phenotypes, highlighting their plasticity. Transcriptional profiling of 9 SCBC and 10 non-SCBC patient tumor samples revealed that SCBC possesses a distinct and unique transcriptional regulatory network. Cell surface proteins associated with the SCBC phenotype showed similarities between PARCB tumors and clinical SCBC tumors. However, the PARCB construct was initially designed to transform prostate/lung cells into prostate/lung NE tumors. This PARCB model does not necessarily recapitulate human NEBC biology, as PARCB is not naturally present in human NEBC tumors. Nevertheless, the results of PARCB transformation support the possibility of the UC-to-NEBC hypothesis.
In a cohort of 34 SCBCs and 84 conventional UCs along with a reference of 408 MIBCs in the TCGA database [37], four groups (Luminal UC, Basal UC, Double-negative UC, and SCBC) were categorized well by unsupervised hierarchical clustering and whole-transcriptome profiles. The SCBC subtype demonstrated significant enrichment of genes associated with promoting cancer metastasis and activating canonical pathways that suppress adaptive immunity. The expression profile change resulted in the lineage plasticity of SCBC, characterized by global downregulation of urothelial differentiation genes and upregulation of NE differentiation programs. The transcriptome analysis data indicated that UC-to-SCBC lineage plasticity was driven by a dysregulated epithelial-to-mesenchymal (EMT) transition combined with a urothelial-to-neural phenotypic switch.
8. Cell of Origin of NEBC
The origin of NEBC has not been definitively determined. Currently, the following hypotheses of cell origin were proposed: 1. pre-existing NE/enterochromaffin cells in the submucosa or normal urothelium; 2. multipotent urothelial stem cell; 3. urinary tract epithelial metaplasia; 4. transformation from UC [30,75]. SCBC closely resembles SCLC in mutation profiles, histological features, and transcriptional patterns. Most lineage tracing studies in SCLC suggest that normal pulmonary NE cells are the cells of origin for SCLC [76]. Therefore, an NE cell origin is generally favored because of the presence of NE cells in the normal urothelium [6]. However, no direct evidence has been reported to confirm that NEBC originates from normal NE cells in the urothelium. The hypothesis of a multipotent stem cell origin is supported by a study showing that microRNA-145 reversed UC in multipotent stem cells, enabling their subsequent differentiation into squamous, NE, and glandular lineages [45]. However, several lines of evidence suggest that NEBC originates from the UC precursor. Most SCBC cases coexist with conventional UC, suggesting that SCBC and UC may share a common clonal origin [63]. This aligns with genetic evidence showing that SCBC and coexisting UC exhibit nearly identical allelic loss patterns [69]. In vitro transduction of five genetic factors (PARCB) successfully transformed human urothelial cells into NEBC tumors, suggesting urothelial cells can be transformed into NEBC cells [74]. Nevertheless, the cell of origin for NEBC remains unclear. Further genetic and molecular studies, along with the development of NEBC preclinical models, are needed to clarify its tumor origin, as this will have significant implications for clinical treatment and management.
9. Potential Therapeutic Targets Identified by Preclinical NEBC Models
Preclinical models are essential for studying tumor progression and evaluating in vitro and in vivo efficacy and safety of potential therapeutics prior clinical trials [77–80]. A NEBC GEMM (Trp53fl/fl Rb1fl/fl MycLSL/LSL) was generated using Lenti-sgNeo#2/Cre delivered via transurethral catheterization, resulting in tumors with SCBC histology [81]. In addition, our lab recently developed another mouse NEBC model (Trp53fl/fl Rb1fl/fl Ptenfl/fl) using Adenovirus-Krt5-Cre. This model produced tumors with NE histology, which were subsequently used to establish stable organoids for drug screening purposes [82]. However, no spontaneous NEBC genetically engineered mouse models (GEMMs) are currently available to study tumor origin, progression, heterogeneity, drug responses, or mechanisms of resistance in NEBC.
Serum-free NEBC cell lines and PDX models were established from two NEBC patients who had not received neoadjuvant treatment [83]. The in vitro growth of NEBC spheroids was found to depend on hepatocyte growth factor (HGF). Treatment with PHA-665752, a MET (MET encoding HGF receptor) inhibitor, significantly reduced NEBC tumor growth. Another PDX (BL100) was developed from a high-grade NEBC on NSG mice [45]. The BL100 PDX model was utilized to evaluate the preclinical efficacy of Rovalpituzumab tesirine (Rova-T), a novel antibody-drug conjugate designed to target DLL3 expression on the surface of SCBC tumor cells by delivering a cytotoxic agent. Rova-T effectively targets DLL3-expressing tumor initiating cells in the BL100 PDX model. Additionally, two of the three SCBC PDXs established in our lab (Figure 2) successfully recapitulated the histology and molecular profiles of SCBC. These PDXs are currently being utilized for in vivo and in vitro drug treatment studies and to evaluate novel therapeutic combinations. These relevant cell lines, GEMMs, and PDX models will serve as valuable tools for studying the tumor origin, progression, heterogeneity, drug response, and resistance in NEBC.
10. Conclusions and Perspectives
NEBCs are rare, accounting for approximately 1% of bladder cancers, with most cases represented as SCBC. SCBC shared some pathogenic features with SCLC but remains distinct in mutational burden driven predominantly by APOBEC-mediated mutational process and diverse epigenetic modifiers [7]. TP53 and RB1 co-mutations are the most common alterations in SCBC. RB1 protein loss is observed in some tumors with the wild-type RB1 gene. Bladder-specific mutations in the TERT promoter and chromatin-modifying genes (e.g., KMT2D, CREBBP, ARID1A, EP300) are also identified in NEBC. ERCC2 mutations, a DNA repair gene, are associated with a complete response to NAC.
NEBC displays significant tumor heterogeneity at the transcriptional level and can be categorized by different classifiers. A unified consensus subtyping system of 6 classifiers for MIBC was recently developed [44], however, many NE-like tumors didn’t exhibit the NE histologically. A single-sample transcriptomic classifier provides a more stringent and accurate definition of the NE-like subtype [43]. Within histologically confirmed NEBC, 4 distinct subsets have been developed by unsupervised hierarchical clustering [45]. Cluster analysis for lineage-specific TFs divided SCBC into three subtypes (ASCL1, NEUROD1, and POU2F3) [38]. This lineage-specific subtyping system was expanded by incorporating the IHC expression patterns of ASCL1, NEUROD1, and POU2F3, resulting in the identification of five distinct subsets [39].
The tumor heterogeneity and plasticity between NEBC and UC haven’t been systematically investigated. The cell of origin of NEBC remains unknown. Recent evidence suggests that SCBC may share a common origin with UC, arising from a UC precursor through UC-to-NEBC plasticity. However, there is also a possibility that NEBC originates from normal NE cells or stem cells within the urothelium. GEMMs of SCLC suggest that both the cell of origin and the genetic driver mutations play a critical role in shaping the phenotypes of SCLC tumors [84]. Hence, GEMMs of SCBC hold promise for providing insights into the cells of origin and identifying the driver lesions and/or epigenetic modifications required for cellular plasticity into NEBC. Additional PDX models will enhance our understanding of NEBC biology and support the development of more effective therapies.
Despite recent advances and collaborative efforts, NEBC continues to present significant challenges and opportunities: 1. identify and validate targetable driver mutations for discovering new therapeutics targets for NEBC; 2. develop more reliable and stable preclinical models (cell line or mouse models) with NEBC features. 3. determine the cell of origin of NEBC with more in vitro and in vivo evidence; 4. understand the mechanism of cell plasticity between UC and NEBC; 5. systemic study and analysis on heterogeneity in NEBC; 6. precisely classify patient samples with NE or NE-like features to select the optimal treatment. Future advances in basic and translational research present promising opportunities to improve outcomes for NEBC patients.
Declarations
Ethics Statement
The study followed the Declaration of Helsinki. Fresh NEBC tumors were obtained from TURBT or cystectomy specimens from de-identified patients at Roswell Park Comprehensive Cancer Center (IRB, BDR# 109219). Written informed consent was obtained from all patients. All animal procedures for PDX generation were approved by the Institutional Animal Care and Use Committee (IACUC) at RPCCC (1395M).
Consent for Publication
Not applicable.
Availability of Data and Material
Not applicable.
Funding
This work was partly supported by NIH Grants, K08CA252161, P30CA016056 (NCI Cancer Center Core Support Grant), the Roswell Park Alliance Foundation, and the Friends of Urology Foundation.
Competing Interests
The authors declare no conflict of interest.
Author Contributions
D.X. and Q.L. wrote and finalized the manuscript.
Acknowledgement
We would like to thank Dr. Bo Xu (RPCCC) for providing H&E and IHC staining for NEBC patient tumors. We thank Dr. David Goodrich and Dr. Dean Tang (RPCCC) for their constructive comments on the manuscript.
Abbreviations
The following abbreviations are used in this manuscript:
- NEBC:
- neuroendocrine bladder cancer
- SCBC:
- small cell bladder cancer
- UC:
- urothelial carcinoma
- SCLC:
- small cell lung cancer
- MIBC:
- muscle-invasive bladder cancer
- WHO:
- World Health Organization
- NE:
- neuroendocrine
- LCNEC:
- large cell neuroendocrine carcinoma
- TURBT:
- transurethral resection of the bladder tumor
- NSE:
- neuron-specific enolase
- INSM1:
- Insulinoma-associated protein 1
- EMA:
- epithelial membrane antigen
- NAC:
- neoadjuvant chemotherapy
- TCGA:
- The Cancer Genome Atlas
- UNC:
- University of North Carolina
- MDA:
- MD Anderson Cancer Center
- H&E:
- hematoxylin and eosin
- IHC:
- immunohistochemistry
- SYP:
- synaptophysin
- CHGA:
- chromogranin-A
- ADC:
- antibody-drug conjugate
- PDX:
- patient-derived xenograft
- TFs:
- transcription factors
- CRISPR:
- clustered regularly interspaced short palindromic repeats
- PARCB:
- a dominant-negative form of TP53 (TP53-DN), myristoylated AKT1 (myr-AKT1), RB1 short-hairpin RNA, C-MYC, and BCL2
- MSKCC:
- Memorial Sloan Kettering Cancer Center
- OS:
- overall survival
- SCCL:
- squamous cell carcinoma-like
- HGF:
- hepatocyte growth factor
- Rova-T:
- Rovalpituzumab tesirine
- GEMMs:
- genetically engineered mouse models
- RPCCC:
- Roswell Park Comprehensive Cancer Center
Table 1. Summary of molecular characterization studies on NEBC.
| Cohort |
Source |
Genomic Seq |
RNA Seq |
Genomic Mutation |
Key Findings |
Citations |
| 307 advanced UC |
Lund University |
N/A |
47 Sc/NE |
N/A |
Lund subtyping taxonomy; mRNA profiling-6 clusters; IHC profiling-5 subtypes; Sc/NE |
2017 Sjödahl et al. [40] |
| 412 MIBC |
TCGA |
20 neuronal |
20 neuronal |
TP53 65%, RB1 25% |
Five subtypes: luminal-papillary, luminal-infiltrated, luminal, basal-squamous, and neuronal |
2017 Robertson et al. [41] |
| 12 SCBC |
Shanghai Jiao Tong University |
12 SCBC |
12 SCBC |
TP53 83%, RB1 33% |
CRISPR in J82 and TCCSUP increased canonical NE markers |
2018 Shen et al. [42] |
| 61 SCBC |
MSKCC |
61 SCBC |
12 SCBC |
TP53 90%, RB1 90%, co-occurred 80%, TERT promoter 95% |
SCBC convergent distinct pathogenesis with SCLC |
2018 Chang et al. [7] |
| PTC, RC1, RC2, NAC, TMT, Imvigor210 |
University of British Columbia |
N/A |
34 NE-like |
N/A |
A strict and accurate single-patient classifier for NE-like |
2019 Batista et al. [43] |
| 1750 MIBC |
Multi-centers |
44 NE-like |
44 NE-like |
NE-like: TP53 94%, RB1 39% (94% present either mutation or deletion) |
Consensus six subtypes: luminal papillary, luminal non-specified, luminal unstable, stroma-rich, basal/squamous, and NE-like |
2019 Kamoun et al. [44] |
| 63 SCBC |
Cleveland Clinic |
N/A |
40 SCBC
6 normal |
N/A |
First subtyping for SCBC, including 4 distinct subsets; DLL3 as a prognostic marker |
2019 Koshkin et al. [45] |
34 SCBC
84 UC |
MD Anderson |
34 SCBC
84 UC |
34 SCBC
84 UC |
TP53 93%, RB1 47%, |
Identical TP53 mutation in paired UC and SCBC; UC-to-NE plasticity drive SCBC |
2020 Yang et al. [37] |
132 SCBC
3236 UC |
Foundation Medicine, Inc. |
132 SCBC |
24 SCBC
51 UC |
TP53 92%, RB1 75%, co-occurred 72%, TERT promoter 68% |
Low T-cell biomarkers; Suppression of inflammatory pathway |
2021 Hoffman et al. [32] |
43 LCNEC
192 SCBC |
MSKCC |
18 LCNEC
52 SCBC |
N/A |
LCNEC: TP53 100%, RB1 100%, co-occurred 56%, TERT promoter 89% |
LNEC longer OS compared with SCBC-only |
2021 Guercio et al. [25] |
| 199 SCBC |
MSKCC |
47 SCBC |
N/A |
TP53 87%, RB1 79%, TERT promoter 83% |
ERCC2 mutation associated with NAC response |
2022 Teo et al. [34] |
| 80 LCNEC |
MSKCC |
37 LCNEC |
N/A |
TP53 92%, RB1 65%, TERT promoter 76% |
Similar genomic profiles of LCNEC and SCBC |
2023 Gandhi et al. [26] |
| 44 SCBC |
Johns Hopkins Medical Institutions |
24 SCBC |
44 SCBC |
TP53 75%, RB1 38%, TERT promoter 92% |
Cluster analysis for lineage-specific TFs (ASCL1, NEUROD1, and POU2F3); high ASCL1 subtype expressed CEACAM5 |
2024 Feng et al. [38] |
103 SCBC
19 LCNBC |
MSKCC |
33 SCBC/LCNBC |
N/A |
TP53 91%, RB1 79%, TERT promoter 82% |
5 subtypes analysis by IHC co-expression pattern of ASCL1, NEUROD1, and POU2F3; DLL3 a therapeutic target |
2024 Akbulut et al. [39] |
References
| 1. |
Siegel RL, Giaquinto AN, Jemal A. Cancer statistics, 2024. CA Cancer J Clin. 2024;74(1):12-49.
[Google Scholar]
[CrossRef]
|
| 2. |
Gandhi J, Chen JF, Al-Ahmadie H. Urothelial Carcinoma: Divergent Differentiation and Morphologic Subtypes. Surg Pathol Clin. 2022;15(4):641-659.
[Google Scholar]
[CrossRef]
|
| 3. |
Kouba E, Cheng L. Neuroendocrine Tumors of the Urinary Bladder According to the 2016 World Health Organization Classification: Molecular and Clinical Characteristics. Endocr Pathol. 2016;27(3):188-199.
[Google Scholar]
[CrossRef]
|
| 4. |
Antoni S, Ferlay J, Soerjomataram I, Znaor A, Jemal A, Bray F. Bladder Cancer Incidence and Mortality: A Global Overview and Recent Trends. Eur Urol. 2017;71(1):96-108.
[Google Scholar]
[CrossRef]
|
| 5. |
Akbulut D, Al-Ahmadie H. Updates on Urinary Bladder Tumors With Neuroendocrine Features. Adv Anat Pathol. 2024;31(3):169-177.
[Google Scholar]
[CrossRef]
|
| 6. |
Cramer SF, Aikawa M, Cebelin M. Neurosecretory granules in small cell invasive carcinoma of the urinary bladder. Cancer. 1981;47(4):724-730.
[Google Scholar]
[CrossRef]
|
| 7. |
Chang MT, Penson A, Desai NB, Socci ND, Shen R, Seshan VE, et al. Small-Cell Carcinomas of the Bladder and Lung Are Characterized by a Convergent but Distinct Pathogenesis. Clin Cancer Res. 2018;24(8):1965-1973.
[Google Scholar]
[CrossRef]
|
| 8. |
Geynisman DM, Handorf E, Wong YN, Doyle J, Plimack ER, Horwitz EM, et al. Advanced small cell carcinoma of the bladder: clinical characteristics, treatment patterns and outcomes in 960 patients and comparison with urothelial carcinoma. Cancer Med. 2016;5(2):192-199.
[Google Scholar]
[CrossRef]
|
| 9. |
Mukesh M, Cook N, Hollingdale AE, Ainsworth NL, Russell SG. Small cell carcinoma of the urinary bladder: a 15-year retrospective review of treatment and survival in the Anglian Cancer Network. BJU Int. 2009;103(6):747-752.
[Google Scholar]
[CrossRef]
|
| 10. |
Thota S, Kistangari G, Daw H, Spiro T. A clinical review of small-cell carcinoma of the urinary bladder. Clin Genitourin Cancer. 2013;11(2):73-77.
[Google Scholar]
[CrossRef]
|
| 11. |
Celik O, Ekin G, Ipekci T, Budak S, Ilbey YO. Diagnosis and treatment in primary bladder small cell carcinoma: Literature review. Arch Ital Urol Androl. 2016;88(1):52-55.
[Google Scholar]
[CrossRef]
|
| 12. |
Fahed E, Hansel DE, Raghavan D, Quinn DI, Dorff TB. Small cell bladder cancer: biology and management. Semin Oncol Nurs. 2012;39(5):615-618.
[Google Scholar]
[CrossRef]
|
| 13. |
Zhao X, Flynn EA. Small cell carcinoma of the urinary bladder: a rare, aggressive neuroendocrine malignancy. Arch Pathol Lab Med. 2012;136(11):1451-1459.
[Google Scholar]
[CrossRef]
|
| 14. |
Koay EJ, Teh BS, Paulino AC, Butler EB. A Surveillance, Epidemiology, and End Results analysis of small cell carcinoma of the bladder: epidemiology, prognostic variables, and treatment trends. Cancer. 2011;117(23):5325-5333.
[Google Scholar]
[CrossRef]
|
| 15. |
Church DN, Bahl A. Clinical review - small cell carcinoma of the bladder. Cancer Treat Rev. 2006;32(8):588-593.
[Google Scholar]
[CrossRef]
|
| 16. |
Lohrisch C, Murray N, Pickles T, Sullivan L. Small cell carcinoma of the bladder: long term outcome with integrated chemoradiation. Cancer. 1999;86(11):2346-2352.
[Google Scholar]
[CrossRef]
|
| 17. |
Choong NW, Quevedo JF, Kaur JS. Small cell carcinoma of the urinary bladder. The Mayo Clinic experience. Cancer. 2005;103(6):1172-1178.
[Google Scholar]
[CrossRef]
|
| 18. |
Abrahams NA, Moran C, Reyes AO, Siefker-Radtke A, Ayala AG. Small cell carcinoma of the bladder: a contemporary clinicopathological study of 51 cases. Histopathology. 2005;46(1):57-63.
[Google Scholar]
[CrossRef]
|
| 19. |
Sved P, Gomez P, Manoharan M, Civantos F, Soloway MS. Small cell carcinoma of the bladder. BJU Int. 2004;94(1):12-17.
[Google Scholar]
[CrossRef]
|
| 20. |
Ismaili N. A rare bladder cancer--small cell carcinoma: review and update. Orphanet J Rare Dis. 2011;6:75.
[Google Scholar]
[CrossRef]
|
| 21. |
Hoffman-Censits J, Choi W, Bivalacqua TJ, Pierorazio P, Kates M, Lombardo K, et al. Small Cell Bladder Cancer Response to Second-line and Beyond Checkpoint Inhibitor Therapy: Retrospective Experience. Clin Genitourin Cancer. 2021;19(2):176-181.
[Google Scholar]
[CrossRef]
|
| 22. |
Pan CX, Yang XJ, Lopez-Beltran A, MacLennan GT, Eble JN, Koch MO, et al. c-kit Expression in small cell carcinoma of the urinary bladder: prognostic and therapeutic implications. Mod Pathol. 2005;18(3):320-323.
[Google Scholar]
[CrossRef]
|
| 23. |
Gupta S, Thompson RH, Boorjian SA, Thapa P, Hernandez LP, Jimenez RE, et al. High grade neuroendocrine carcinoma of the urinary bladder treated by radical cystectomy: a series of small cell, mixed neuroendocrine and large cell neuroendocrine carcinoma. Pathology. 2015;47(6):533-542.
[Google Scholar]
[CrossRef]
|
| 24. |
Wang G, Yuan R, Zhou C, Guo C, Villamil C, Hayes M, et al. Urinary Large Cell Neuroendocrine Carcinoma: A Clinicopathologic Analysis of 22 Cases. Am J Surg Pathol. 2021;45(10):1399-1408.
[Google Scholar]
[CrossRef]
|
| 25. |
Guercio BJ, Gandhi J, Teo MY, Lattanzi M, Funt SA, Aggen DH, et al. Large cell neuroendocrine carcinoma of the urothelial tract (LNEC): The MSKCC experience. J Clin Oncol. 2021;39 15_suppl:4526.
[Google Scholar]
[CrossRef]
|
| 26. |
Gandhi JGB, Mirsadraei L, Zhou Q, Chen JF, Sarungbam J, Chen YB, et al. Pathological, clinical and molecular characterization of large cell neuroendocrine carcinoma (LCNEC) of the urinary tract. Lab Invest. 2023;103 3, Supplement:S727-39.
[Google Scholar]
|
| 27. |
Chen JF, Yang C, Sun Y, Cao D. Expression of novel neuroendocrine marker insulinoma-associated protein 1 (INSM1) in genitourinary high-grade neuroendocrine carcinomas: An immunohistochemical study with specificity analysis and comparison to chromogranin, synaptophysin, and CD56. Pathol. Res Pract. 2020;216(6):152993.
[Google Scholar]
[CrossRef]
|
| 28. |
Kim IE Jr., Amin A, Wang LJ, Cheng L, Perrino CM. Insulinoma-associated Protein 1 (INSM1) Expression in Small Cell Neuroendocrine Carcinoma of the Urinary Tract. Appl Immunohistochem Mol Morphol. 2020;28(9):687-693.
[Google Scholar]
[CrossRef]
|
| 29. |
Hoang LL, Tacha D, Bremer RE, Haas TS, Cheng L. Uroplakin II (UPII), GATA3, and p40 are Highly Sensitive Markers for the Differential Diagnosis of Invasive Urothelial Carcinoma. Appl Immunohistochem Mol Morphol. 2015;23(10):711-716.
[Google Scholar]
[CrossRef]
|
| 30. |
Sanguedolce F, Calò B, Chirico M, Tortorella S, Carrieri G, Cormio L. Urinary Tract Large Cell Neuroendocrine Carcinoma: Diagnostic, Prognostic and Therapeutic Issues. Anticancer Res. 2020;40(5):2439-2447.
[Google Scholar]
[CrossRef]
|
| 31. |
George J, Lim JS, Jang SJ, Cun Y, Ozretić L, Kong G, et al. Comprehensive genomic profiles of small cell lung cancer. Nature. 2015;524(7563):47-53.
[Google Scholar]
[CrossRef]
|
| 32. |
Hoffman-Censits J, Choi W, Pal S, Trabulsi E, Kelly WK, Hahn NM, et al. Urothelial Cancers with Small Cell Variant Histology Have Confirmed High Tumor Mutational Burden, Frequent TP53 and RB Mutations, and a Unique Gene Expression Profile. Eur Urol Oncol. 2021;4(2):297-300.
[Google Scholar]
[CrossRef]
|
| 33. |
Zheng X, Zhuge J, Bezerra SM, Faraj SF, Munari E, Fallon JT 3rd, et al. High frequency of TERT promoter mutation in small cell carcinoma of bladder, but not in small cell carcinoma of other origins. J Hematol Oncol. 2014;7:47.
[Google Scholar]
[CrossRef]
|
| 34. |
Teo MY, Guercio BJ, Arora A, Hao X, Regazzi AM, Donahue T, et al. Long-term Outcomes of Local and Metastatic Small Cell Carcinoma of the Urinary Bladder and Genomic Analysis of Patients Treated With Neoadjuvant Chemotherapy. Clin Genitourin Cancer. 2022;20(5):431-441.
[Google Scholar]
[CrossRef]
|
| 35. |
van Allen EM, Mouw KW, Kim P, Iyer G, Wagle N, Al-Ahmadie H, et al. Somatic ERCC2 mutations correlate with cisplatin sensitivity in muscle-invasive urothelial carcinoma. Cancer Discov. 2014;4(10):1140-1153.
[Google Scholar]
[CrossRef]
|
| 36. |
Liu D, Plimack ER, Hoffman-Censits J, Garraway LA, Bellmunt J, Van Allen E, et al. Clinical Validation of Chemotherapy Response Biomarker ERCC2 in Muscle-Invasive Urothelial Bladder Carcinoma. JAMA Oncol. 2016;2(8):1094-1096.
[Google Scholar]
[CrossRef]
|
| 37. |
Yang G, Bondaruk J, Cogdell D, Wang Z, Lee S, Lee JG, et al. Urothelial-to-Neural Plasticity Drives Progression to Small Cell Bladder Cancer. iScience. 2020;23(6):101201.
[Google Scholar]
[CrossRef]
|
| 38. |
Feng M, Matoso A, Epstein G, Fong M, Park YH, Gabrielson A, et al. Identification of Lineage-specific Transcriptional Factor-defined Molecular Subtypes in Small Cell Bladder Cancer. Eur Urol. 2024;85(6):523-526.
[Google Scholar]
[CrossRef]
|
| 39. |
Akbulut D, Whiting K, Teo MY, Tallman JE, Ozcan GG, Basar M, et al. Differential NEUROD1, ASCL1, and POU2F3 Expression Defines Molecular Subsets of Bladder Small Cell/Neuroendocrine Carcinoma With Prognostic Implications. Mod Pathol. 2024;37(10):100557.
[Google Scholar]
[CrossRef]
|
| 40. |
Sjödahl G, Eriksson P, Liedberg F, Höglund M. Molecular classification of urothelial carcinoma: global mRNA classification versus tumour-cell phenotype classification. J Pathol. 2017;242(1):113-125.
[Google Scholar]
[CrossRef]
|
| 41. |
Robertson AG, Kim J, Al-Ahmadie H, Bellmunt J, Guo G, Cherniack AD, et al. Comprehensive Molecular Characterization of Muscle-Invasive Bladder Cancer. Cell. 2017;171(3):540-556.e25.
[Google Scholar]
[CrossRef]
|
| 42. |
Shen P, Jing Y, Zhang R, Cai MC, Ma P, Chen H, et al. Comprehensive genomic profiling of neuroendocrine bladder cancer pinpoints molecular origin and potential therapeutics. Oncogene. 2018;37(22):3039-3044.
[Google Scholar]
[CrossRef]
|
| 43. |
Batista da Costa J, Gibb EA, Bivalacqua TJ, Liu Y, Oo HZ, Miyamoto DT, et al. Molecular Characterization of Neuroendocrine-like Bladder Cancer. Clin Cancer Res. 2019;25(13):3908-39020.
[Google Scholar]
[CrossRef]
|
| 44. |
Kamoun A, de Reyniès A, Allory Y, Sjödahl G, Robertson AG, Seiler R, et al. A Consensus Molecular Classification of Muscle-invasive Bladder Cancer. Eur Urol. 2020;77(4):420-433.
[Google Scholar]
[CrossRef]
|
| 45. |
Koshkin VS, Garcia JA, Reynolds J, Elson P, Magi-Galluzzi C, McKenney JK, et al. Transcriptomic and Protein Analysis of Small-cell Bladder Cancer (SCBC) Identifies Prognostic Biomarkers and DLL3 as a Relevant Therapeutic Target. Clin Cancer Res. 2019;25(1):210-221.
[Google Scholar]
[CrossRef]
|
| 46. |
Lindgren D, Frigyesi A, Gudjonsson S, Sjödahl G, Hallden C, Chebil G, et al. Combined gene expression and genomic profiling define two intrinsic molecular subtypes of urothelial carcinoma and gene signatures for molecular grading and outcome. Cancer Res. 2010;70(9):3463-3472.
[Google Scholar]
[CrossRef]
|
| 47. |
Damrauer JS, Hoadley KA, Chism DD, Fan C, Tiganelli CJ, Wobker SE, et al. Intrinsic subtypes of high-grade bladder cancer reflect the hallmarks of breast cancer biology. Proc Natl Acad Sci U S A. 2014;111(8):3110-3115.
[Google Scholar]
[CrossRef]
|
| 48. |
Sjödahl G, Lauss M, Lövgren K, Chebil G, Gudjonsson S, Veerla S, et al. A molecular taxonomy for urothelial carcinoma. Clin Cancer Res. 2012;18(12):3377-3386.
[Google Scholar]
[CrossRef]
|
| 49. |
Choi W, Porten S, Kim S, Willis D, Plimack ER, Hoffman-Censits J, et al. Identification of distinct basal and luminal subtypes of muscle-invasive bladder cancer with different sensitivities to frontline chemotherapy. Cancer Cell. 2014;25(2):152-165.
[Google Scholar]
[CrossRef]
|
| 50. |
Weinstein JN, Akbani R, Broom BM, Wang W, Verhaak RGW, McConkey D, et al. Comprehensive molecular characterization of urothelial bladder carcinoma. Nature. 2014;507(7492):315-322.
[Google Scholar]
[CrossRef]
|
| 51. |
Marzouka NA, Eriksson P, Rovira C, Liedberg F, Sjödahl G, Höglund M. A validation and extended description of the Lund taxonomy for urothelial carcinoma using the TCGA cohort. Sci Rep. 2018;8(1):3737.
[Google Scholar]
[CrossRef]
|
| 52. |
Grivas P, Bismar TA, Alva AS, Huang HC, Liu Y, Seiler R, et al. Validation of a neuroendocrine-like classifier confirms poor outcomes in patients with bladder cancer treated with cisplatin-based neoadjuvant chemotherapy. Urol Oncol. 2020;38(4):262-268.
[Google Scholar]
[CrossRef]
|
| 53. |
Rudin CM, Poirier JT, Byers LA, Dive C, Dowlati A, George J, et al. Molecular subtypes of small cell lung cancer: a synthesis of human and mouse model data. Nat Rev Cancer. 2019;19(5):289-297.
[Google Scholar]
[CrossRef]
|
| 54. |
Baine MK, Hsieh MS, Lai WV, Egger JV, Jungbluth AA, Daneshbod Y, et al. SCLC Subtypes Defined by ASCL1, NEUROD1, POU2F3, and YAP1: A Comprehensive Immunohistochemical and Histopathologic Characterization. J Thorac Oncol. 2020;15(12):1823-1835.
[Google Scholar]
[CrossRef]
|
| 55. |
Chan JM, Quintanal-Villalonga Á, Gao VR, Xie Y, Allaj V, Chaudhary O, et al. Signatures of plasticity, metastasis, and immunosuppression in an atlas of human small cell lung cancer. Cancer Cell. 2021;39(11):1479-1496.e18.
[Google Scholar]
[CrossRef]
|
| 56. |
Kang HW, Kim WJ, Choi W, Yun SJ. Tumor heterogeneity in muscle-invasive bladder cancer. Transl Androl Urol. 2020;9(6):2866-2880.
[Google Scholar]
[CrossRef]
|
| 57. |
Tang DG. Understanding and targeting prostate cancer cell heterogeneity and plasticity. Semin Cancer Biol. 2022;82:68-93.
[Google Scholar]
[CrossRef]
|
| 58. |
Tang DG, Kondo T. Cancer cell heterogeneity and plasticity: From molecular understanding to therapeutic targeting. Semin Cancer Biol. 2022;82:1-2.
[Google Scholar]
[CrossRef]
|
| 59. |
McGranahan N, Swanton C. Clonal Heterogeneity and Tumor Evolution: Past, Present, and the Future. Cell. 2017;168(4):613-628.
[Google Scholar]
[CrossRef]
|
| 60. |
Meeks JJ, Al-Ahmadie H, Faltas BM, Taylor JA 3rd, Flaig TW, DeGraff DJ, et al. Genomic heterogeneity in bladder cancer: challenges and possible solutions to improve outcomes. Nat Rev Urol. 2020;17(5):259-270.
[Google Scholar]
[CrossRef]
|
| 61. |
Dyrskjøt L, Hansel DE, Efstathiou JA, Knowles MA, Galsky MD, Teoh J, et al. Bladder cancer. Nat Rev Dis Primers. 2023;9(1):58.
[Google Scholar]
[CrossRef]
|
| 62. |
Wang G, Xiao L, Zhang M, Kamat AM, Siefker-Radtke A, Dinney CP, et al. Small cell carcinoma of the urinary bladder: a clinicopathological and immunohistochemical analysis of 81 cases. Hum Pathol. 2018;79:57-65.
[Google Scholar]
[CrossRef]
|
| 63. |
Blomjous CE, Vos W, De Voogt HJ, Van der Valk P, Meijer CJ. Small cell carcinoma of the urinary bladder. A clinicopathologic, morphometric, immunohistochemical, and ultrastructural study of 18 cases. Cancer. 1989;64(6):1347-1357.
[Google Scholar]
[CrossRef]
|
| 64. |
Shenoy S. Cell plasticity in cancer: A complex interplay of genetic, epigenetic mechanisms and tumor micro-environment. Surg Oncol. 2020;34:154-162.
[Google Scholar]
[CrossRef]
|
| 65. |
Yuan S, Norgard RJ, Stanger BZ. Cellular Plasticity in Cancer. Cancer Discov. 2019;9(7):837-851.
[Google Scholar]
[CrossRef]
|
| 66. |
Ku SY, Wang Y, Garcia MM, Yamada Y, Mizuno K, Long MD, et al. Notch signaling suppresses neuroendocrine differentiation and alters the immune microenvironment in advanced prostate cancer. J Clin Invest. 2024;134(17):e175217.
[Google Scholar]
[CrossRef]
|
| 67. |
Gui Y, Guo G, Huang Y, Hu X, Tang A, Gao S, et al. Frequent mutations of chromatin remodeling genes in transitional cell carcinoma of the bladder. Nat Genet. 2011;43(9):875-878.
[Google Scholar]
[CrossRef]
|
| 68. |
Comprehensive molecular characterization of urothelial bladder carcinoma. Nature. 2014;507(7492):315-322.
[Google Scholar]
[CrossRef]
|
| 69. |
Cheng L, Jones TD, McCarthy RP, Eble JN, Wang M, MacLennan GT, et al. Molecular genetic evidence for a common clonal origin of urinary bladder small cell carcinoma and coexisting urothelial carcinoma. Am J Pathol. 2005;166(5):1533-1539.
[Google Scholar]
[CrossRef]
|
| 70. |
Priemer DS, Wang M, Zhang S, Lopez-Beltran A, Kouba E, Montironi R, et al. Small-cell Carcinomas of the Urinary Bladder and Prostate: TERT Promoter Mutation Status Differentiates Sites of Malignancy and Provides Evidence of Common Clonality Between Small-cell Carcinoma of the Urinary Bladder and Urothelial Carcinoma. Eur Urol Focus. 2018;4(6):880-888.
[Google Scholar]
[CrossRef]
|
| 71. |
Niederst MJ, Sequist LV, Poirier JT, Mermel CH, Lockerman EL, Garcia AR, et al. RB loss in resistant EGFR mutant lung adenocarcinomas that transform to small-cell lung cancer. Nat Commun. 2015;6(1):6377.
[Google Scholar]
[CrossRef]
|
| 72. |
Ku SY, Rosario S, Wang Y, Mu P, Seshadri M, Goodrich ZW, et al. Rb1 and Trp53 cooperate to suppress prostate cancer lineage plasticity, metastasis, and antiandrogen resistance. Science. 2017;355(6320):78-83.
[Google Scholar]
[CrossRef]
|
| 73. |
Mu P, Zhang Z, Benelli M, Karthaus WR, Hoover E, Chen CC, et al. SOX2 promotes lineage plasticity and antiandrogen resistance in TP53- and RB1-deficient prostate cancer. Science. 2017;355(6320):84-88.
[Google Scholar]
[CrossRef]
|
| 74. |
Wang L, Smith BA, Balanis NG, Tsai BL, Nguyen K, Cheng MW, et al. A genetically defined disease model reveals that urothelial cells can initiate divergent bladder cancer phenotypes. Proc Natl Acad Sci U S A. 2020;117(1):563-572.
[Google Scholar]
[CrossRef]
|
| 75. |
Kouba EJ, Cheng L. Understanding the Genetic Landscape of Small Cell Carcinoma of the Urinary Bladder and Implications for Diagnosis, Prognosis, and Treatment: A Review. JAMA Oncol. 2017;3(11):1570-8.
[Google Scholar]
[CrossRef]
|
| 76. |
Park KS, Liang MC, Raiser DM, Zamponi R, Roach RR, Curtis SJ, et al. Characterization of the cell of origin for small cell lung cancer. Cell Cycle. 2011;10(16):2806-2815.
[Google Scholar]
[CrossRef]
|
| 77. |
Guo H, Xu X, Zhang J, Du Y, Yang X, He Z, et al. The Pivotal Role of Preclinical Animal Models in Anti-Cancer Drug Discovery and Personalized Cancer Therapy Strategies. Pharmaceuticals (Basel). 2024;17(8):1048.
[Google Scholar]
[CrossRef]
|
| 78. |
Ireson CR, Alavijeh MS, Palmer AM, Fowler ER, Jones HJ. The role of mouse tumour models in the discovery and development of anticancer drugs. British Journal of Cancer. 2019;121(2):101-108.
[Google Scholar]
[CrossRef]
|
| 79. |
Sajjad H, Imtiaz S, Noor T, Siddiqui YH, Sajjad A, Zia M. Cancer models in preclinical research: A chronicle review of advancement in effective cancer research. Animal Model Exp Med. 2021;4(2):87-103.
[Google Scholar]
[CrossRef]
|
| 80. |
Golebiewska A, Fields RC. Advancing preclinical cancer models to assess clinically relevant outcomes. BMC cancer. 2023;23(1):230.
[Google Scholar]
[CrossRef]
|
| 81. |
Liao R, Imamura J, Muskara A, Ganguly S, Balyimez A, Ornstein M, et al. A NOVEL ORTHOTOPIC MURINE MODEL OF NEUROENDOCRINE BLADDER CANCER: INSIGHTS INTO THE MOLECULAR DRIVERS OF SMALL CELL BLADDER CANCER (SCBC). Urologic Oncology: Seminars and Original Investigations. 2024;42:S31.
[Google Scholar]
[CrossRef]
|
| 82. |
Xu D, Cao Q, Wang L, Wang Y, Zhang X, Wu Y, et al. Abstract B13: A novel mouse model for small-cell carcinoma of bladder. Cancer Res. 2020;80 11_Supplement:B13.
[Google Scholar]
[CrossRef]
|
| 83. |
Hofner T, Macher-Goeppinger S, Klein C, Rigo-Watermeier T, Eisen C, Pahernik S, et al. Development and characteristics of preclinical experimental models for the research of rare neuroendocrine bladder cancer. J Urol. 2013;190(6):2263-2270.
[Google Scholar]
[CrossRef]
|
| 84. |
Ferone G, Lee MC, Sage J, Berns A. Cells of origin of lung cancers: lessons from mouse studies. Gene Dev. 2020;34(15–16):1017-1032.
[Google Scholar]
[CrossRef]
|


 ,
Qiang Li
1,2
,
Qiang Li
1,2